受精
受精(fertilization)是精子与卵子融合形成受精卵的过程。成熟的精子和发育中的卵子(处于减数分裂Ⅱ中期的次级卵母细胞)如果不发生受精,将会在几分钟或数小时内死亡。受精挽救了精子和卵子,标志着一个新生命的开始。
一、受精的条件
在哺乳动物和人类,受精一般发生在输卵管壶腹部。因此,作为受精的条件,除取决于精子和卵子(次级卵母细胞)的发育成熟之外,性交后进入雌性生殖管道的精子必须获得活力,即游动与受精的能力。
完成减数分裂变形阶段的精子脱离睾丸的支持细胞,进入曲细精管的管腔中,而后开始迁移,依次通过直细精管、睾网、输出小管进入附睾管和输精管,在附睾尾和输精管内储存、成熟,直至射精。精子在附睾内受其分泌的甘油磷酸胆碱、内毒碱、唾液酸糖蛋白等作用下,逐渐达到功能上的成熟。
研究发现,哺乳动物的精子进入雌性生殖管道后并不立即受精,而是停留一段时间,以获得对卵子受精的能力,这一过程称为精子的获能(capacitation)。在人类,精子的获能需要5~6小时。获能后的精子表现为游动和细胞呼吸能力的增加,它们像卫星一样围绕着卵子细胞游动,其运动力的增加提高了精子与卵子细胞相互作用而接近的概率。
精子获能的本质是:精子因雌性生殖管道的“特殊条件”而被改变。较多的实验证据表明,在精子进入雌性生殖管道后,先由于某种未知的原因造成精子内部的钾离子外流,引起质膜静息电位发生变化,增加了质膜的不稳定性,致使膜胆固醇脱落,促使雌性生殖管道中的HCO3ˉ和Ca2+进入精子细胞,继而激活了质膜上的腺苷酸环化酶,使精子细胞内cAMP水平升高,通过PKA激活胞内蛋白酪氨酸激酶,促使多种蛋白质产生磷酸化,从而提高了精子的代谢和运动能力,并使细胞膜处于超极化状态。
获能也改变了精子细胞膜的脂质和糖蛋白的组分。例如,精液中含有唾液酸糖蛋白,并附着在精子头部的外表面,阻止其顶体酶(acrosomal enzyme)的释放;当精子在子宫和输卵管中运动时,该糖蛋白被此处分泌物中唾液酸酶α和β淀粉酶等降解,使精子获得授精能力。
精子的数量和质量是保证受精的条件之一。性交后射出的3亿个人精子中,只有200个能够到达输卵管的受精地点。有证据表明,排卵周围的卵泡细胞释放化学信号来吸引精子到达卵细胞,但这种化学信号的本质尚不清楚。一次射精的精液中,精子总数大于4×107,其中能向前运动的大于50%,精子形态正常的大于30%。受精虽然只由一个精子和一个卵子完成,但因精子与卵子的接触有一定的随机性,所以需要较多的精子到达输卵管壶腹部,才能在限定时间内有正常精子完成与卵细胞的结合。
二、受精过程
在哺乳动物和人类,性交后,当获能的精子与发育中的卵子(处于减数分裂Ⅱ中期的次级卵母细胞)相遇时,将引发系列连锁反应,包括:精子识别卵细胞并诱发顶体反应,精子穿过卵外被抵达卵细胞膜,精-卵质膜融合后精子细胞核等成分进入卵细胞,次级卵母细胞完成减数分裂,皮层反应阻止多精入卵,以及精-卵核融合等事件(图14-7)。精子与卵子的融合,标志着一个新生命的开始。
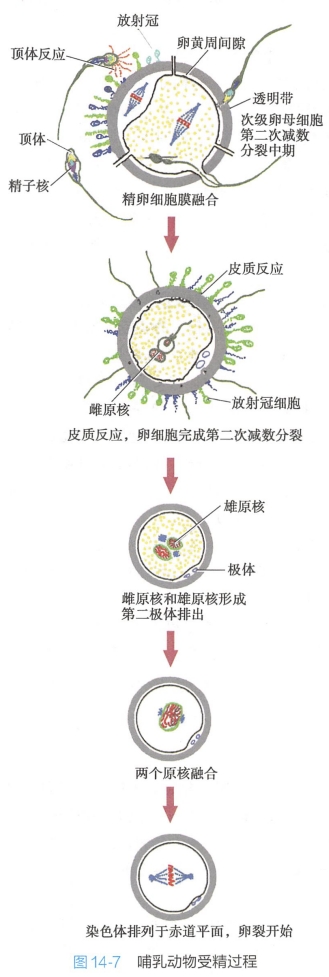
(一)精子识别卵细胞并穿过卵外被
1.精子卵子识别诱发顶体反应
精子一旦获能,便穿过卵泡细胞层,结合到透明带。透明带是受精的种属屏障,去除透明带就等于消除了这个屏障。例如,如果将大鼠卵子的透明带经酶消化去除后,人类的精子就可以使其受精,当然,这样的杂合子受精卵无法继续发育。
哺乳动物卵子的透明带主要由ZP1、ZP2和ZP3这3种糖蛋白质组成,它们均由生长中的卵母细胞产生。3种ZP蛋白在透明带中的排列不是随机的,ZP2和ZP3聚集成长丝状,而ZP1则将这些长丝交联在一起成为三维的网络结构。在这三种蛋白中,ZP1仅具有结构功能,而ZP2和ZP3则同时参与配子间的相互作用。
- 一般认为ZP3是精子与透明带结合的主要受体,可以诱导顶体反应(acrosome re- action)。ZP3受体的特异性由其肽链上的O-连接寡糖所决定。ZP3缺陷会导致雌性小鼠不育。精子与卵细胞透明带上的ZP3受体结合后,将触发Ca2+向精子细胞质的内流,进而引发顶体反应,即精子的顶体成分通过胞吐作用被释放出来,所释放的多种水解酶可以消化卵子的透明带,以帮助精子穿过透明带;顶体反应是一种特殊的细胞胞吐过程。研究表明,精子中顶体的系列变化自获能的精子接触放射冠时即开始。获能的精子首先与卵子周围的放射冠接触。这时精子顶体的前膜即与表面的细胞膜融合,继而破裂形成许多小孔,顶体内含的酶(酸性水解酶)逐渐释放出来。当精子与透明带上的ZP3受体结合后,导致顶体破裂,内含物完全释放。在精子接触透明带之前,卵丘细胞分泌的孕酮启动顶体反应,而精子与透明带结合后,启动顶体反应的是ZP3。如此,顶体反应中释放的顶体酶先解离放射冠的卵泡细胞,继而分解透明带,形成一个精子穿过的通道。
- ZP2可作为经过顶体反应的精子的次级受体(secondary receptor)。顶体反应也把精子中某些能与ZP2结合的蛋白质释放到精子的表面,使精子在进入卵子过程中与透明带紧密连接。在顶体反应期间,与ZP3结合的顶体前端发生胞吐作用,致使精子表面与ZP3结合的配体蛋白随之丢失,精子必须继续与透明带结合才能完成穿透过程,此时是通过顶体内膜上的特殊蛋白与透明带中的ZP2糖蛋白结合而实现的。这表明,顶体反应后精子与透明带的结合由ZP3转到ZP2,ZP2作为精子的次级受体进一步加固精子与透明带的结合。
2.精子穿过透明带抵达卵细胞膜
在卵子的受精过程中,精子以种特异性的方式结合到透明带上,从而完成顶体反应,穿过透明带到达质膜,精子与卵子细胞膜的直接接触,标志受精的开始。
(二)精-卵融合
1.精-卵质膜融合
精子细胞核等成分进入卵细胞受精开始时,精子头侧面的细胞膜与卵细胞膜融合,随即精子的细胞核和细胞质进入卵内。
卵子细胞膜表面覆盖有许多的微绒毛,精子首先作用于微绒毛顶端的细胞质膜,邻近的微绒毛随即迅速地伸长、聚丛,包绕着精子,从而保证精子与卵子紧密结合并发生质膜融合。当精子质膜与卵子质膜融合时,微绒毛被吸收,精子以头部先入的方式进入卵子细胞质中。
精-卵质膜融合的机制至今尚不清楚。迄今人们在精子中的ADAM(a disintegrin and metalloprotease)家族成员和卵子细胞膜上的整联蛋白(integrin)在精-卵质膜融合中的作用,进行了较多研究。小鼠精子细胞膜上存在有ADAM家族成员一受精素(fertilin),它是在顶体反应后暴露于精子细胞表面的跨膜蛋白,能够帮助精子结合到卵子质膜并促进精-卵质膜融合。
受精素由两个糖基化的跨膜亚单位α链和β链组成,它们之间以非共价键结合。受精素α亚单位的胞外部分含有一个疏水区,它在结构上类似于介导病毒与感染细胞融合的病毒融合蛋白,该区域的人工合成多肽能够诱导试管内的精-卵质膜融合。然而一些体外实验发现,受精素缺失的精子仍然可以使卵子受精,这提示尚存在其他精子蛋白参与到精卵质膜融合过程。
受精素的细胞外N-端区域能够同卵子质膜上的整联蛋白(α6β1)结合,使精子黏附在卵子的表面以便为精-卵融合做准备。
2.次级卵母细胞完成减数分裂过程
精子进入卵子后,激发次级卵母细胞迅速完成第二次减数分裂,形成一个成熟的卵子和一个第二极体,卵子细胞单倍染色体向中央移行,核膜形成,形成雌原核(female pronucleus)。精子进入卵后,核膜崩遗,尾部退化消失,细胞染色质解聚,核内精蛋白被组蛋白替换,新形成的原核膜包在染色质外周,形成雄原核(male pronucleus)。
3.精-卵膜融合后发生皮层反应阻止多精入卵
虽然有许多精子可以与卵子结合,但通常只有一个精子能够与卵子的胞膜融合并向卵子细胞质内释放出它的细胞核和其他细胞器。如果有多余一个的精子与卵子融合,则称为多精入卵(polyspermy),此时多极或过多的纺锤体形成,将导致细胞分裂时染色体的错误分配、非二倍体细胞的产生以及发育的停滞。
迄今研究表明,有两个调控机制可以确保只有一个精子与卵子结合。
- 一是卵子与第一个精子融合后将引起卵子质膜的快速去极化,以阻止其他精子与已受精卵子的融合,这是一个快速的早期阻止多精入卵的机制。
- 但卵子质膜的极性在受精作用后很快恢复正常,因此需要另外一个作用时间长的机制来阻止多精入卵。这就是卵子的皮层反应(cortical reaction),即卵子通过胞吐作用将其皮质颗粒释放出来,改变卵外被结构。
卵子细胞中储存的Ca2+的释放与皮层反应直接相关。精子与卵子的质膜融合,将激活胞内内质网上存在的Ca2+通道,引起储存Ca2+的释放,导致卵子局部细胞质内游离Ca2+浓度的增加,并形成钙波(calcium wave)传播到整个卵子,启动皮层反应。
哺乳动物的研究表明,皮层颗粒中的糖苷酶降解透明带中的ZP3为ZP3f,ZP3f缺少糖基,致使精子膜表面蛋白无法识别;皮层颗粒中的蛋白酶降解ZP2为ZP2f,ZP2f不能与顶体反应精子结合,使多余的精子不能进入,从而阻止多精入卵。
在卵子发生过程中,透明带蛋白是泛素化的。用泛素蛋白酶体抑制剂处理卵子,可阻止精子进入卵子的透明带,表明去泛素化也可阻止多精入卵。
(三)精-卵核融合完成受精过程
卵子一旦受精后即为合子。只有当两个单倍体的细胞核(雄原核和雌原核)融合在一起,并且它们的染色体“混合”成为一个二倍体的细胞核时,受精过程才算完成。
在哺乳动物,受精卵内的两个原核并不像其他物种那样直接融合,它们彼此接近但是保持距离,这种状态一直持续到合子细胞准备进行第一次有丝分裂(即卵裂)时为止。此时,在卵子胞质中的微管和微丝作用下,雌原核、雄原核相互靠近,接触处原核膜呈指状,相互交错对插;同时染色体浓缩,原核膜崩解、消失,精-卵染色体组合在一起,形成合子染色体组,定位于纺锤体上。
此时细胞即为受精卵,受精至此完成。
多数动物(包括人类)在受精时,精子为合子提供的不仅仅是DNA,它还提供中心粒(在人类未受精的卵子里不存在)。精子中心粒伴随着细胞核和尾端进入卵子,并在其周围形成中心体。在人类,精子中心粒进行复制并且组织合子细胞第一次有丝分裂的纺锤体装配。这也解释了在多精入卵时为什么有多极现象和多余的纺锤体形成(由于多个精子的中心粒进入卵子)。
受精具有重要意义:
- 受精使卵子的缓慢代谢转入代谢旺盛期,从而启动细胞不断地分裂;
- 精子与卵子的结合,恢复了二倍体,维持物种的稳定性;
- 受精决定性别,带有Y染色体的精子与卵子结合发育为男性,带有X染色体的精子与卵子结合则发育为女性;
- 受精的染色体来自父母双方,加之生殖细胞在成熟分裂时曾发生染色体联会和片断交换,使遗传物质重新组合,使新个体具有与亲代不完全相同的性状。
三、辅助生殖技术
辅助生殖技术(assisted reproductive technolog,ART)。ART是指应用现代生物医学知识、技术及方法对配子、合子及胚胎进行人工操作,代替人类自然生殖过程中的某些步骤,以达到受孕目的的一项技术,主要包括体外受精、胚胎移植及其衍生技术。迄今辅助生殖技术已催生出四代试管婴儿:
(一)第一代试管婴儿
体外受精(in vitro fertilization,IVF)技术产生了世界上第一例试管婴儿。体外受精是指分别将卵子与精子取出后,置于试管内使其受精,经过数天的生长后(卵裂期或囊胚期),再将胚胎移植(embryo transfer,ET)到母体子宫内发育成胎儿并分娩的技术,该技术统称为常规IVF-ET,最终分娩的婴儿称为试管婴儿(第一代试管婴儿)。该成果在2010年获得诺贝尔医学奖。IVF-ET的受精率只有30%~40%。
IVF-ET的适用范围是:
- 因女性因素所致不孕问题,如各种原因导致的输卵管不通或功能障碍(输卵管切除、输卵管阻塞、盆腔严重粘连及输卵管结扎再通术失败等);
- 男方少、弱精子症;
- 不明原因的不育;
- 免疫性不孕等。
(二)第二代试管婴儿
卵胞质内单精子注射(intracytoplasmic sperm injection,ICSI)技术,即借助显微操作系统将单一精子注射人卵子内使其受精。ICSI技术可以解决常规VF-ET成功率不高的问题,受精率可达50%~70%,提高了VF的成功率。ICSI技术仅需数个精子就可以达到受精、妊娠,是严重男性因素不育患者的最有效治疗方法。它是适用于男性少、弱精子症患者,以及圆头(顶体缺乏)精子或完全不活动精子等。ICSI的成功率很大程度上受精子质量和显微操作技术的影响。ICSI潜在的问题可能是精子的表观遗传“程度”不够。
ICSI和IVF是男性不育症的最有效治疗方法,婴儿活产率为20%~25%。
(三)第三代试管婴儿
胚胎移植前遗传学诊断(preimplantation genetic diagnosis,PGD)技术是用于解决人类遗传病而发展的一项技术。在受精卵分裂到6~8个细胞的时候,对透明带打孔后,取出一个细胞进行PGD,选择正常的胚胎植入子宫,避免父母有遗传性疾病的遗传缺陷婴儿的出生,达到优生的目的。
PGD技术主要解决的是:
- 夫妻中一人可能携带了遗传病基因或异常染色体,想确定自己的后代将来是否会受到该基因的影响;
- 夫妻双方想要确定,自己是否携带了异常染色体以至于不能正常怀孕。
(四)第四代试管婴儿
有些女性虽然有排卵功能,但是身体条件不好,或年龄偏大,致使卵子质量不高、活力差、线粒体异常等,不适合使用上述技术。这时可输入另一位健康女性卵细胞质形成一个新的优质卵细胞,将此新的卵子与其丈夫精子结合成受精卵,植入子宫内,妊娠分娩。此技术称为卵浆置换(exchange of cytoplasmic and nucleus)技术(第四代试管婴儿)。
目前,世界上已普遍开展了精子和胚胎的冷冻和复融技术,而卵子的冷冻和复融成功率较低,正在探索中。国内现在大部分VF实验室也相继建立了精子及胚胎冻融技术。 ART有利于解决不孕症,但也引发许多伦理问题,如生育与性行为的分离,传统家庭模式的解体,亲子关系的破裂,代孕母亲的利弊,精子库、卵子库的功过、胚胎的地位等等。