生殖细胞
结构简单的生物,如原核生物或单细胞真核生物,通常进行无性生殖(asexual reproduction);结构复杂的高等动物和植物,一般都进行有性生殖(sexual reproduction)。
在有性生殖过程中,必须有两个亲本参加,先形成配子(gamete),例如哺乳动物中的精子(spermatozoa或sperm)和卵子(ovum或egg)。精子与卵子结合后形成合子(zygote)或称受精卵(fertilized ovum或fertilized egg),由受精卵通过有丝分裂发育成下一代新个体。
配子是由生殖系细胞(germ line)产生的。通常把有性生殖过程中专门供两性细胞融合的精子和卵子称为成熟生殖细胞。精子和卵子均为单倍体(haploid)细胞,它们融合后所形成的受精卵为二倍体(diploid)细胞。例如,人类的配子仅有23条染色体,其中一条是性染色体(X或Y染色体),受精后成为46条染色体(男性性染色体为XY;女性性染色体为XX)。
一、生殖细胞的起源
在哺乳动物,如小鼠受精后,受精卵经卵裂(cleavage)、桑葚胚(morula)、囊胚(blastocyst)发育形成具有三胚层的原肠胚(gastrula)。
囊胚由两层不同的细胞构成,分别为上胚层(epiblast)和上胚层内面的内细胞团(inner cell mass)。在三胚层形成之前,上胚层中的祖细胞(progenitor cell)群区域化,产生原始生殖细胞(primordial germ cell,PGC)。PGC的产生需要产生自内脏内胚层(visceral endoderm)与胚外外胚层(extraembryonic ectoderm)的TGF-β和Wnt等信号转导途径的参与。产生的PGC迁移到生殖嵴(genital ridge)(即未来的性腺)(图14-2)。 #组胚
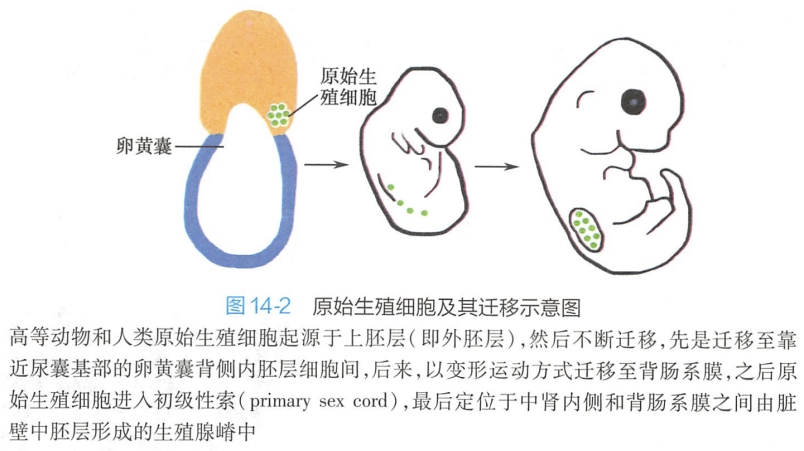
在生殖嵴处,PGC分化成为生殖干细胞(germ stem cell)。生殖干细胞提供能用于生殖的配子。PGC的迁移和增殖受到周围体细胞分泌的许多因子(如PRDM1和PRDM14)的控制。 PGC迁移到生殖嵴时,性别决定(sex determination)即开始。人类性分化大致开始于孕后第6周。通常认为分化成为卵巢是默认途径,因为XY型生殖嵴分化成为睾丸是在SRY(sex-determining region on the chromosome Y)基因控制下进行的,而XX型生殖嵴没有SRY基因,因而发育成卵巢。一旦PGC迁移到生殖腺嵴中,性腺即开始发育,性腺的发育受到体细胞产生的许多转录因子(如GATA4、 FOXL2、LHX9、WT1、WNT4和SF1)的调控。
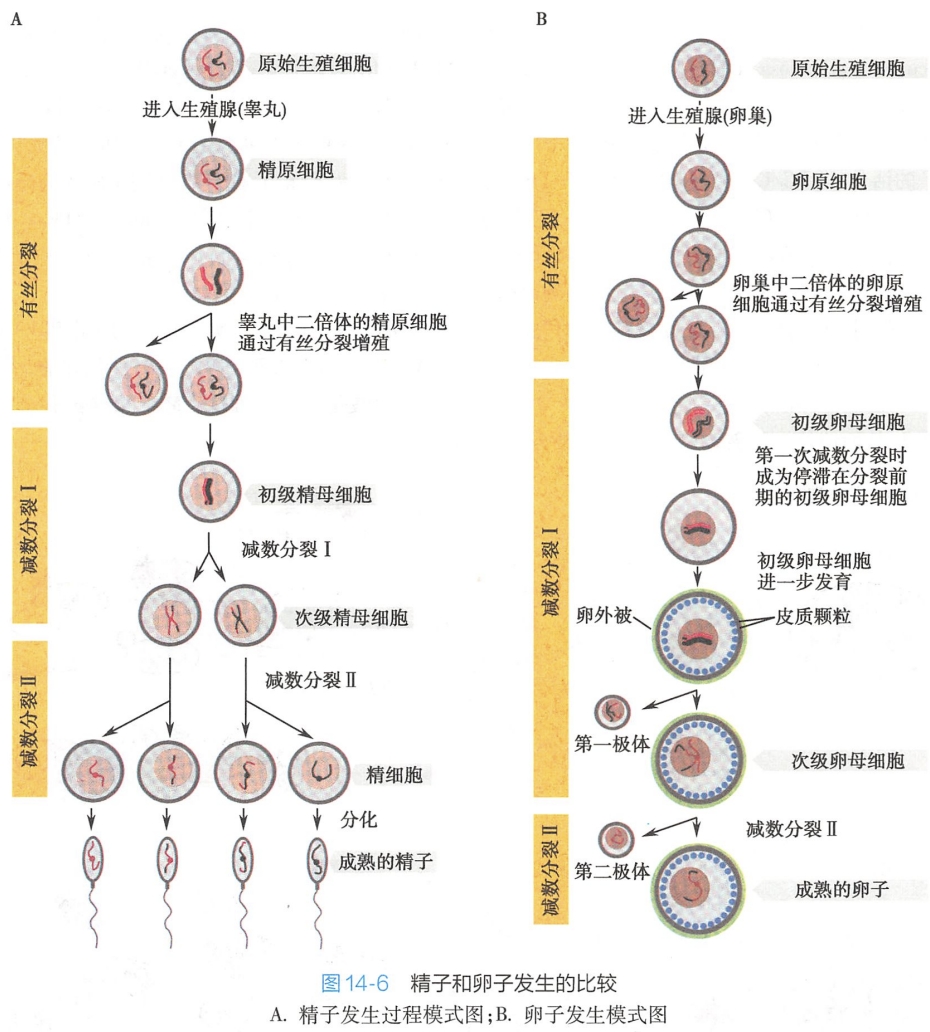
在哺乳动物,位于胚胎性腺的原始生殖细胞通过有丝分裂增殖并分化为生殖细胞,生殖细胞经历了两种类型的分裂,即有丝分裂和减数分裂,最终分化为成熟的配子(精子和卵子)。精子和卵子的形成,称为配子发生(gametogenesis),是指二倍体的生殖细胞发育成单倍体的精子或卵子的过程。也可分别称为精子发生(spermatogenesis)和卵子发生(oogenesis)。
二、精子的发生
(一)精子由精原细胞发育而来
在胎儿期,睾丸中的生殖细胞并不进入减数分裂,而是停留在G0/G1期,直到出生后才继续增殖,其中一部分迁移到睾丸的生精小管的基底膜(basement membrane),并转变为精原细胞(spermatogonium)或精原干细胞(spermatogonial stem cell)。
精原细胞可以通过有丝分裂增殖,并通过减数分裂产生精细胞(spermatid)。这些精细胞通过核凝缩、胞质的减少、顶体和鞭毛的形成,而在功能和形态上成为精子。
人类精子的发生开始于青春期,产生于睾丸的生精小管(即曲细精管),然后迁移到附睾中成熟并排出。
生精小管是一种特殊的复层上皮管道,称这种上皮为生精上皮(germinal epithelium或ser miniferous epithelium)。生精上皮包含两种类型的生发细胞(germinal cell),一种是形成精子的生精细胞,自生精小管外表面至腔面分别分布精原细胞、初级精母细胞、次级精母细胞、精子细胞和精子;另一种是起支持、免疫、营养及分泌等功能的支持细胞(sertoli cell),为精子产生所必需。精原细胞发育为精子的过程称为精子发生,在人类约为64天,其发生过程可分为增殖期、生长期、成熟期和变形期4个时期。
1.增殖期为精原细胞的有丝分裂
精原细胞位于基膜上,呈圆形,分化程度较低。青春期后,精原细胞开始不断增殖,可分为A、B两型。A型精原细胞不断增殖,一部分保留下来作为干细胞,稳定精原细胞数量和保持活跃的生精能力;另一部分分化为B型精原细胞,可生长分化为精子。精原细胞细胞核中染色体数目为2倍体(2n),人精原细胞为46条染色体。
2.生长期的本质是初级精母细胞的形成与发育
B型精原细胞体积增大,形成初级精母细胞,其染色体数目仍为2n。初级精母细胞处于第一次减数分裂期,随着染色质的变化,可分为细线期(leptotene)、偶线期(zygotene)及粗线期(pachytene)精母细胞。粗线期精母细胞的体积可达到细线期的两倍以上。第一次减数分裂的时间较长,在人可达22天,分裂后形成次级精母细胞。
3.成熟期为精母细胞的减数分裂期
初级精母细胞形成后,迅速进入减数分裂I(即第一次分裂),形成2个次级精母细胞(secondary spermatocyte)。次级精母细胞存在的时间较短,很快完成减数分裂Ⅱ(即第二次分裂),最终形成4个精细胞。上述两次分裂,只在第一次分裂中染色体复制一次,而细胞却分裂两次,结果形成的4个精子细胞中染色体数目都减少一半,由2n变为n(参见图14-6A)。人的精细胞有23条染色体(n)。
4.变形期的本质是精细胞特化为成熟的精子
精细胞位于生精小管的近腔面,体积更小,细胞呈圆形(称为圆形精子,round spermatid),细胞不再分裂,经复杂的变态发育后形成精子(elongating spermatid)(图14-3)。
精子包括两个形态与功能不同的区域,即头和鞭毛(头和鞭毛连接处特称为颈部),这些区域均为质膜所包裹。头部(head)含有凝缩的单倍体细胞核,被称为顶体(acrosome)的囊泡所包裹,顶体来自高尔基复合体,含有穿透卵外被的蛋白水解酶;鞭毛(flagellum)区域又可细分为中段、主段和端部。中段(body)含有中心粒,富含线粒体,中心粒可作为产生鞭毛的基体,而线粒体产生鞭毛运动所需要的能量。
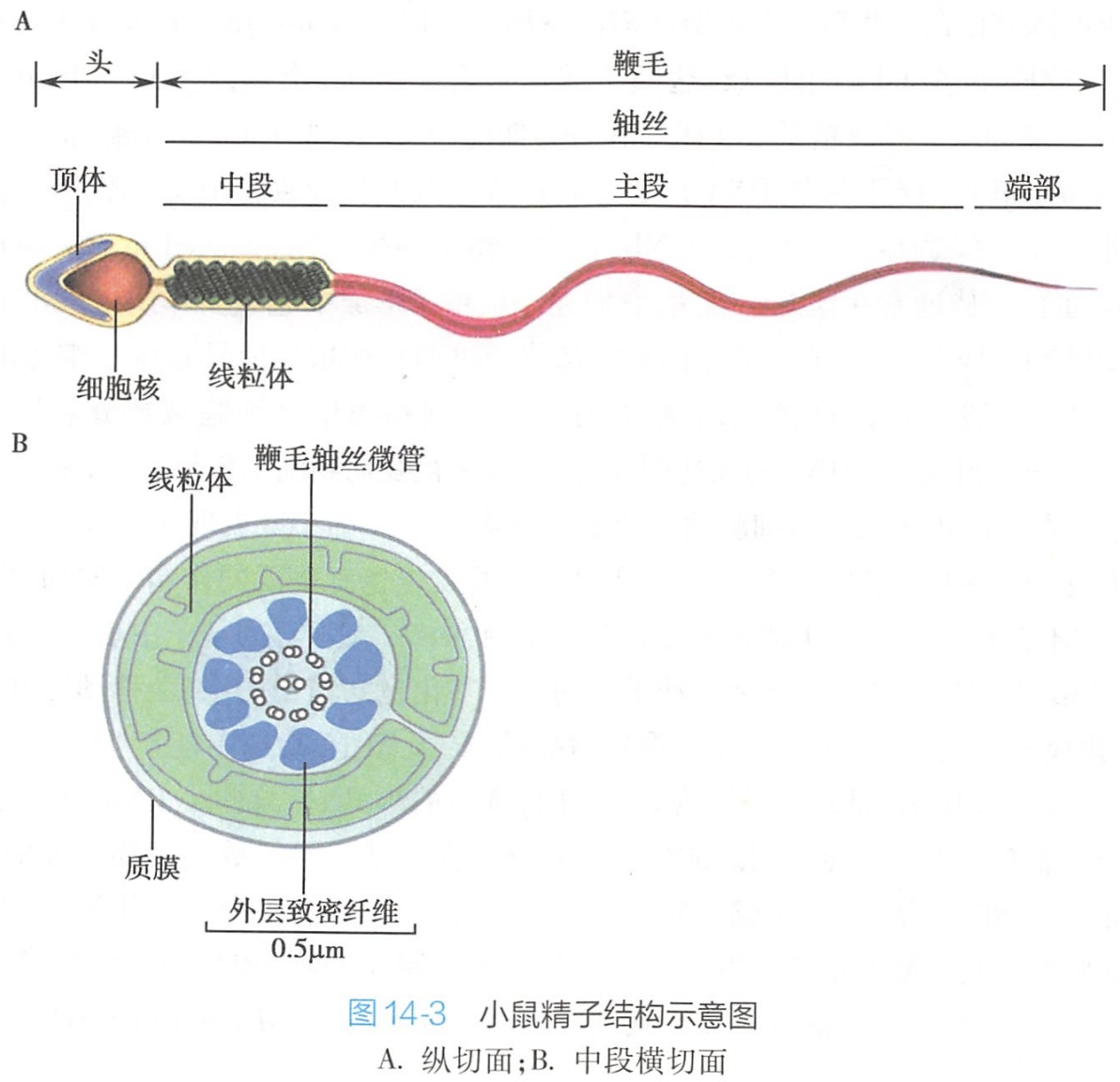
(1)顶体形成
精细胞的高尔基复合体经过变化形成一个囊泡,称为顶体囊泡。顶体囊泡与核膜相粘并增大,变为半球状覆盖于核前面,不断增大呈帽状,套于核前2/3形成顶体(acrosome)。
顶体中含有透明质酸酶、酸性磷酸酶、神经氨酸苷酶及蛋白酶等。顶体具有类似溶酶体的功能,在受精时顶体释放这些酶,消化卵表面的透明带,形成通道,有助于精子穿过透明带进入卵子细胞内。
(2)中心粒的变化
中心粒移至顶体的对侧,近侧中心粒保留,远侧中心粒形成轴丝,颈部的外面由漏斗状结构包裹,漏斗的扩大部分固着于核的尾端,而另一端则与精子尾部的中段相连。在轴丝的外侧出现与之平行的9条外周致密纤维,构成主段的纤维鞘。哺乳类动物的远侧中心粒一般在身部形成后将消失。
(3)线粒体改变
精子细胞的线粒体体积变小并伸长,且迁移到终端,围绕着中央轴丝形成螺旋状排列的线粒体鞘。线粒体鞘为鞭毛运动提供能量,使精子得以快速运动。
(4)细胞核的特化
精子细胞由圆形变为长形,细胞核变为扁平梨形。精子生成过程的早期, DNA与组蛋白结合,但是随后大部分精子中的组蛋白被精蛋白(protamine)所取代,使得染色质以一种特殊的方式包装,浓缩形成精子头部。精蛋白与DNA的结合也抑制了DNA的表达,增加了抗机械损伤的能力。
精子在生成过程中,组蛋白被替代是逐步进行的,首先是睾丸特异的组蛋白变异体取代正常的组蛋白,然后转变蛋白(transition protein)取代组蛋白变异体。最后,精蛋白取代了转变蛋白。受精后,精子中的精蛋白被卵子中的组蛋白重新取代。精蛋白或转变蛋白的缺陷会导致小鼠精子不育或增加不育性。
(5)尾部形成
构成尾部全长轴心是轴丝,由“9+2”排列的微管组成,来自位于细胞核后端的基体(basal body),随细胞变长相应伸长。细胞质向尾部汇集并脱落,精子细胞从圆形变为蝌蚪状精子。
(二)多种因子影响精子的发生
一般而言,因素可分为外源性的和内源性的。外源性因素主要包括内分泌、旁分泌和自分泌因子的调控,而内源性因素主要是指生精细胞内基因水平的调控。
1.分泌因子的调控
支持细胞、睾丸间质(Leydig)细胞、生殖细胞以及周围的其他细胞分泌的因子都参与了精子的发生。睾丸生成睾酮(testosterone)和精子。下丘脑和垂体以及睾丸本身所产生的激素调节睾酮和精子的产生。
下丘脑分泌的促性腺素释放因子(gonadotropin-releasing hormone,GnRH)通过垂体刺激促黄体素(luteinizing hormone,LH)和促卵泡激素(follicle stimulating hormone,FSH)的产生,LH通过血液运输到睾丸,并在睾丸处刺激间质细胞产生睾酮(作为雄激素),睾酮也可环化雌激素。
睾酮也可反馈下丘脑和垂体,抑制GnRH等的产生。
雄激素和FSH也可以对支持细胞产生效应,形成最适的精子发生环境。 #生理
2.基因的调控
从分子水平上来看,精子的发生和成熟分化过程是一系列特定基因程序性表达的结果。参与调控的有转录因子基因、细胞周期相关基因、原癌基因、调亡及自噬相关基因、热休克蛋白基因、细胞骨架蛋白基因、核蛋白转型相关基因等。涉及基因的转录调控、前体RNA剪切及翻译水平的调控、蛋白质的稳定性调控(如泛素化)等。人类Y染色体上存在一些精子发生所必需的基因,如AZF(azoospermia factor)DNA序列。
又如,Pwi蛋白家族通常在睾丸中表达,它们在小鼠精子形成中起重要作用。Pwi蛋白家族成员包括MILI、MIWI和MIWI2,任一成员基因的剔除都将通过凋亡的增加而导致生殖细胞的丧失。
DNA甲基化、miRNA和组蛋白-精蛋白转变对精子的发生也有重要的作用。
又如 piRNA(Piwi-interacting RNA)是一类长度为24~32核苷酸的小RNA,这些小RNA只有与Piwi蛋白家族成员相结合才能发挥它的调控作用。piRNA主要分布在包括人类等数种动物睾丸的精原细胞内,它们在减数分裂开始时大量积聚,在成熟的精子产物中消失。研究结果提示,piRNA与Piwi共同作用调控生殖干细胞的自我更新及精子的发育成熟。其主要机制是Piwi-piRNA复合物引起基因沉默、维持基因组稳定。
3.环境因子影响精子的发生
通常环境因子分为下列类型:雌激素类似物、二氧(杂)芑(二噁英)、邻苯二甲酸盐、多氯化联(二)苯、氟化合物、多环芳烃、溴系阻燃剂以及重金属。
三、卵子的发生
(一)卵子由卵原细胞发育而来、直至受精后才发育成熟
卵原细胞(oogonium)发育为卵子的过程称为卵子发生,一般可分为增殖期、生长期、成熟期三个时期。
1.增殖期的特点是有丝分裂和卵原细胞的形成
PGC在生殖嵴中,经过多次有丝分裂,成为卵原细胞。卵原细胞通过有丝分裂迅速增殖,在人体中可达到5百万~7百万个细胞,但是其中大部分卵原细胞将会死亡,仅留下约5万个。
2.生长期是初级卵母细胞的再发育
卵原细胞体积增大并进一步发育成初级卵母细胞(primary oocyte),在这个时期(通常为哺乳动物出生以前),减数分裂I开始。此后,初级卵母细胞停留在减数分裂I的分裂前期(前期I的双线期),此期可长达数月(小鼠)或数十年(女性)。
在绝大多数哺乳类动物中,LH可诱导卵细胞的最后成熟。此后,初级卵母细胞进一步发育,在细胞内积累大量卵黄、RNA和蛋白质等物质,为受精后的发育提供信息、物质和能量的准备,其染色体数仍为二倍体(2n)。雌性生殖细胞是在卵泡(follicle)中发育成熟的。初级卵母细胞在原始卵泡和生长卵泡内进一步发育。
(1)原始卵泡
原始卵泡(primordial follicle)位于卵巢皮质浅层,体积小,数量多。卵泡中央为初级卵母细胞,周围为单层扁平的卵泡细胞(follicle cell),卵泡细胞具有支持和营养卵母细胞的作用。卵泡细胞与卵母细胞之间有许多缝隙连接,有利于卵泡细胞将营养物质输送给卵母细胞以及进行细胞间离子、激素和小分子物质的交换。
(2)生长卵泡
原始卵泡生长发育转变为生长卵泡(growing follicle),卵泡逐渐移至卵巢皮质深层,主要变化是初级卵母细胞体积增大,卵泡细胞和卵泡周围结缔组织增生,生长卵泡可分为初级卵泡和次级卵泡两个时期:
- 初级卵泡(primary follicle),由原始卵泡发育形成。此时初级卵母细胞体积增大,卵泡细胞由单层扁平状变为立方形或柱状,随之细胞增殖成多层(5~6层),卵泡周围间质细胞密集形成卵泡膜。在初级卵泡早期,卵母细胞和卵泡细胞之间出现一层含糖蛋白的透明带(zona pellucida)(图14-4)。
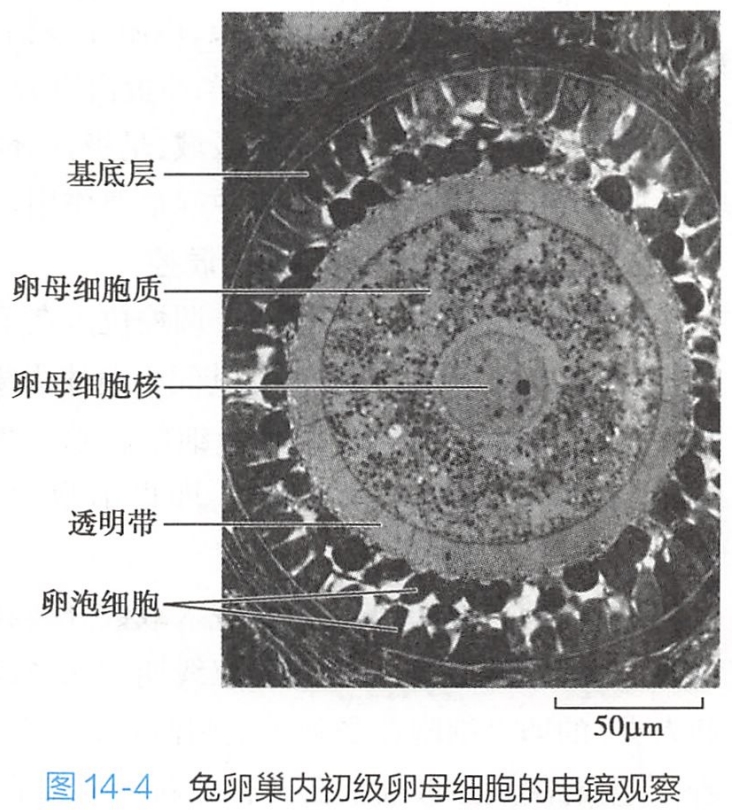
透明带由卵泡细胞和初级卵母细胞共同分泌形成的,具有较强的组织特异性和抗原性,能对同种精子进行专一性的识别与结合,从而使受精过程具有相当的物种专一性。透明带也能保护卵子免于机械损伤。初级卵母细胞还在此时产生含有用于受精的酶和蛋白质的皮质颗粒(cortical granule)。皮质颗粒含有含蛋白酶、黏多糖等。 - 初级卵泡继续生长成为次级卵泡(sc ondary follicle)。此时卵泡体积更大,卵泡细胞增至 6~12层,细胞间出现一些不规则的腔隙,并逐渐合并成一个半月形的腔,称为卵泡腔(follicullar an- tum),腔内充满卵泡液,内含透明质酸酶和性激素及营养物质成分。卵泡液是由卵泡细胞分泌液和卵泡膜血管渗出液组成的。
- 随着卵泡液的增多及卵泡腔扩大,卵母细胞居于卵泡的一侧,并与其周围的颗粒细胞一起突向卵泡腔,形成卵丘(cumulus oophorus),此时的卵泡称为近成熟卵泡(premature follicle)。紧贴透明带的一层柱状卵泡细胞呈放射状排列,称放射冠(corona radiata);分布在卵泡腔周边的卵泡细胞较小,构成卵泡壁,称为颗粒层(stratum granulosum)。在卵泡生长过程中,卵泡膜分化为内、外两层。卵泡的生长主要受LH和FSH的影响。 #组胚
3.成熟期的标志是次级卵母细胞的形成
当卵泡增大至直径约15~20mm时向卵巢表面突出,即为成熟卵泡(mature follicle)。成熟卵泡的卵泡腔很大,颗粒层甚薄,颗粒细胞也不再增殖。成熟卵泡由卵丘细胞、透明带、卵周腔和卵子细胞组成。
成熟卵泡是卵泡发育的最后阶段,通常是在性成熟时期发生的。发育成熟的初级卵母细胞在激素的影响下,细胞开始完成减数分裂I,染色体重新凝缩,核被膜裂解,复制的同源染色体在后期I分离,成为两个子细胞核,每个都包含了原有染色体的一半,但是细胞质不对称分裂产生两个在大小上有很大不同的细胞:一个是小的极体(polar body)(即第一极体),另一个是大的次级卵母细胞。
在此阶段,每个染色体仍然是由两个姐妹染色单体构成。在大多数哺乳类动物中,次级卵母细胞紧接着进入减数分裂Ⅱ,并被阻断在减数分裂Ⅱ的中期,只有在受精后才最终完成减数分裂。人卵巢中的初级卵母细胞大约只有400个能进入成熟期,发育为排卵时停止在减数分裂Ⅱ的中期的次级卵母细胞。
排卵(ovulation)是指突出于卵巢表面的成熟卵泡发生破裂,包含有卵丘细胞的卵母细胞随卵泡液排除的过程。每月有1个卵细胞成熟并被排出。
4.次级卵母细胞在受精后完成减数分裂Ⅱ而形成成熟卵子
受精后,次级卵母细胞的姐妹染色单体在减数分裂Ⅱ后期分离之后,大的次级卵母细胞的细胞质再次发生不对称分裂产生成熟的卵子和一个小的第二极体。极体与卵子一样,也是单倍体。因在卵细胞发育全过程中,细胞质发生了两次不对称分裂,使卵细胞保持了较大的体积。所形成的三个极体(第一极体仍可分裂为2个极体)体积很小,最终将退化消失(图14-6B)。发育成熟的卵子细胞具有显著特征:
- 在其质膜外有一层卵外被(egg coat),为一种特殊形式的细胞外基质,主要由大量的糖蛋白分子构成。在哺乳动物卵外被即为透明带:非哺乳动物为卵黄膜。卵外被可以保护卵细胞免受机械损伤,同时又是受精过程中精子的种属特异性屏障,它只允许物种相近或相同的精子进入卵子细胞。
- 包括哺乳动物在内,许多卵子细胞在其质膜下靠近胞质外侧的区域为皮层(cortex)(其实就是细胞皮层吧),内含高浓度的肌动蛋白分子和内含特化的分泌囊泡,即皮层颗粒(cortical granule)。皮层颗粒包含有消化酶、黏多糖、黏性糖蛋白和透明蛋白,当卵子被精子激活时,这些皮层颗粒成分通过胞吐作用(exocytosis)被释放出来,改变卵外被(在哺乳动物为透明带;非哺乳动物为卵黄膜)的性质,以阻止多精入卵。
- 成熟卵子中合成了大量的蛋白质、核糖体,以及tRNA和mRNA等,某些mRNA和蛋白质为指导受精卵发育的形态发生因子(morphogenetic factor),它们通常定位于卵细胞的不同区域,呈极性分布,在卵裂过程中被分离到不同的细胞中去,这些成分在胚胎极性的确定及模式形成中起重要作用。
(二)卵母细胞减数分裂的调控
卵母细胞减数分裂的第一个调控位点在第一次减数分裂双线期。卵母细胞可以在该期停留很长时间(在人类可以达到50年),其间染色体去凝聚并进行活跃的转录,转录活性升高可表现为卵母细胞体积明显增大。例如,人卵母细胞在这一时期的直径可达到100μm。此期中,细胞生长的同时,卵母细胞蓄积大量RNA和蛋白质等支持胚胎早期发育所需要的物质。
不同物种的卵母细胞重新进入减数分裂和受精的发生过程是不同的。某些动物的卵母细胞在受精前一直停留在减数分裂I的双线期,在受精后才完成减数分裂。但是,大多数脊椎动物(包括蛙、鼠和人类)的卵母细胞在受到激素刺激后发生减数分裂的恢复,并在受精前完成第一次减数分裂。接着,产生的次级卵母细胞在没有重新形成细胞核,染色体也没有去凝聚的情况下进入第二次减数分裂。然后,大多数脊椎动物卵母细胞又会停滞在减数分裂Ⅱ中期,并维持在这个时期直到受精。
卵母细胞停止在前期I是由于MPF活性低造成的,恢复减数分裂需要MPF的活化。MPF由 p34Cdc2激酶(CDK1或Cdc2)亚基和周期蛋白B1所构成。CDK1在G2/M期转变过程中也是必需的。几种信号转导途径能确保停止在前期I的卵母细胞中的MPF处于低活性状态,然后再活化MPF重新进入减数分裂(图14-5A):
1.LH低
(1)cAMP-PKA
在卵母细胞停留在前期I时,壁颗粒细胞(mural granulosa cell)产生Nppc,Nppc刺激卵丘颗粒细胞(cumulus granulosa cell)产生鸟苷酸环化酶NPr2,NPr2催化GTP生成cGMP。cGMP通过Cx37缝隙连接进入卵母细胞,防止了PDE3A在卵母细胞中水解cAMP成为5'-腺苷酸。cAMP活化PKA,
- PKA转过来活化Wee1 B激酶,Wee1 B激酶磷酸化CDK1,导致了CDK1的钝化,即MPF的钝化。
- PKA同时抑制Cdc25B磷酸酶,阻止Cdc25B去磷酸CDK1,从而阻止MPF活化。
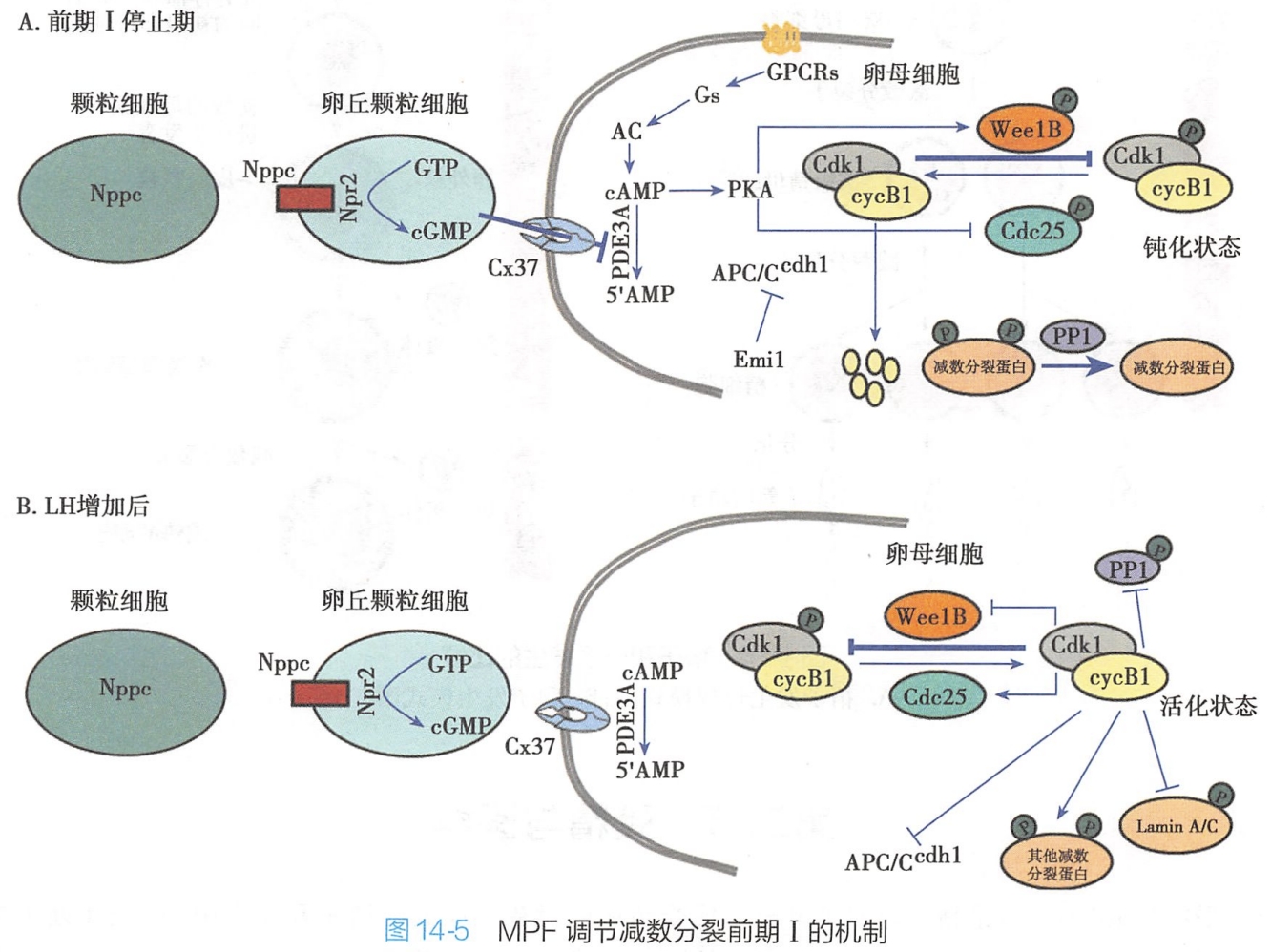
(2)APC/Ccdh1
APC/Ccdh1介导周期蛋白B1(cyelin B1)连续降解,阻止了MPF在前期I停止的卵母细胞中的活化。APC/C是一个多亚基的E3泛素连接酶,正向调节蛋白Cdh1是APC/C活化和底物特异性所必需)。前期I停止的卵母细胞中APC/Ccdh1可被Emi1蛋白抑制。
(3)PP1
低MPF活性也有利于PP1的活化,PP1可连续地去除减数分裂蛋白的磷酸化。
2.LH增加
排卵时LH的增加导致了缝隙连接的关闭,阻止了cGMP进入卵母细胞。从而增加了PDE3A介导的cAMP的水解。低水平的cAMP和PKA不再活化Wee1 B和磷酸化并钝化Cdc25B,CDK1去磷酸化并活化。
活化的MPF钝化APC/Ccdh1。
活化的MPF磷酸化并钝化PP1。这有利于其他CDK1底物处于磷酸化状态的维持。
MPF可磷酸化核纤层蛋白A,从而使核被膜破裂,继续进行减数分裂(图14-5B)。
四、哺乳动物的精子和卵子在发生上的差异
尽管精子和卵子的发生过程都经历减数分裂,但在大多数哺乳动物,精子和卵子的产生过程有很大不同,主要表现在:
- 在发生与成熟的时间上不同,例如,在人类女性,卵原细胞仅在胎儿时期增殖,出生以前就进入了减数分裂I期,并且停留在卵母细胞状态,可以持续50多年。卵细胞按照严格的发育程序成熟并定期排卵,基本上从青春期开始第一次排卵,而且排卵后的次级卵母细胞的核仍是二倍的,直到受精后减数分裂才完成;与女性相比,男性精原细胞的减数分裂和精子的形成直到青春期才在睾丸中开始。
- 在形成成熟生殖细胞的数量上不同,精子发生中的减数分裂,细胞为对称分裂,产生四个相同的精子细胞;而卵子发生的减数分裂是不对称的,形成一个卵子和三个极体(图14-6)。