细胞分裂
细胞分裂(cell division)是细胞生命活动的重要特征之一,指一个亲代细胞形成两个子代细胞的过程。
通过细胞分裂,亲代细胞的遗传物质和某些细胞组分可以相对均等地分配到两个子代细胞中,这有效地保证了生物遗传的稳定性。
在单细胞生物(如细菌、酵母等)中,细胞分裂是其个体繁衍的重要方式,在高等生物中,经过多次细胞分裂生成成熟的性细胞,这些性细胞之间的有性生殖导致受精卵产生,新的生命个体由此出现。
细胞分裂构成了多细胞生物个体生长的基础,从受精开始到个体成熟的整个发育进程中,细胞需经历多次分裂,最终形成机体的器官和组织中数量庞大的细胞群体。
细胞分裂在维持和更新个体正常组织中也具有重要的作用,在成体动物的皮肤、骨髓、肠上皮等器官组织中存在一些具有潜在分裂能力的原始细胞,如表皮基底层和毛囊中的干细胞、骨髓造血干细胞和肠上皮干细胞等,通过这些细胞的分裂可以适时地生成大量的新的分化成熟细胞,以替代因生理性衰老而死亡的细胞。
此外,在机体创伤后的组织修复和再生等活动中都存在大量的细胞分裂现象。细胞分裂的过程总是呈周期性进行:亲代分裂产生子代细胞;子代细胞经历一系列规律的细胞内生物化学变化,包括遗传物质的复制和特定蛋白质的合成等准备过程,并伴有细胞形态学的改变;然后子代细胞的分裂过程开始。
通常将细胞从上次分裂结束到下次分裂结束所经历的规律性变化过程称为一个细胞周期(cell cycle)。大多数的细胞周期都包括数个协调的过程:细胞生长、DNA复制、倍增的染色体分配到子细胞中及细胞分裂。
在细菌,细胞生长和DNA复制发生在细胞周期的多数时间内,倍增的DNA和质膜一起分配到子细胞中。在真核细胞,细胞周期比较复杂,包含4个分离的时相期。虽然细胞生长通常是一个连续的过程,但DNA合成仅仅发生在细胞周期的一个时相中,然后,复制的染色体通过一个连续的过程在细胞分裂前被分配到子细胞核中。
细胞分裂的方式主要包括有丝分裂、减数分裂及无丝分裂三种,不同分裂方式在分裂过程和子代细胞的遗传特性等方面各具特点。
一、有丝分裂
有丝分裂(mitosis)也称间接分裂(indirect division),是高等真核生物的体细胞分裂的主要方式。
有丝分裂是细胞分裂的一系列事件连续发生和发展的过程。有丝分裂持续时间约0.5~2小时,是一个连续的动态变化过程,包括细胞核分裂和胞质分裂,也是形态学变化最为丰富的时期。
根据分裂细胞的形态和结构的变化,以细胞核分裂为坐标,即从细胞分裂开始到细胞核分裂的过程,通常人为地将有丝分裂划分为五个时期:前期;前中期:中期;后期;末期。
此外,胞质分裂期则可从有丝分裂的后期启动,延续至末期。通过核分裂及胞质分裂两个过程,借助于细胞骨架的重排,有丝分裂的细胞实现了染色体及胞质在子代细胞中的均等分配。
染色质凝集、纺锤体及收缩环的形成是有丝分裂活动中的三个重要的特征。
蛋白质磷酸化与去磷酸化是有丝分裂中染色质凝集与去凝集、核膜解聚与重建等变化产生的分子基础(图13-1)。
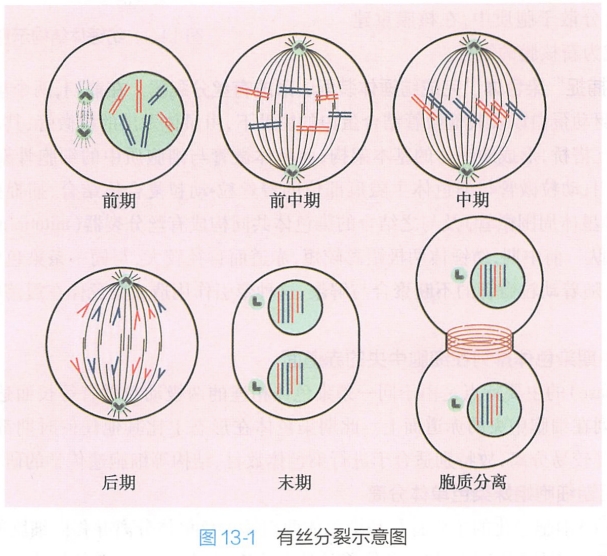
(一)分裂前期核内染色质开始凝集
前期(prophase)细胞变化的主要特征为:染色质凝集、分裂极确定、纺锤体形成、核仁缩小解体。
1.染色质凝集成染色体
伴随核仁缩小、消失,间期核松散染色质纤维螺旋化并发生折叠,导致染色质纤维凝集变粗变短是细胞进入有丝分裂前期的标志。在染色质凝集过程中,因染色质上的核仁组织中心组装到了所属染色体中,导致rRNA合成停止,核仁开始逐渐分解,并最终消失。
2.分裂极确立和纺锤体的形成
随着染色质的凝集,原来分布于细胞同一侧并已经完成复制的两个中心体(centrosome)开始沿核膜外围分别向细胞的两极移动,它们最后到达的位置将决定细胞分裂极。
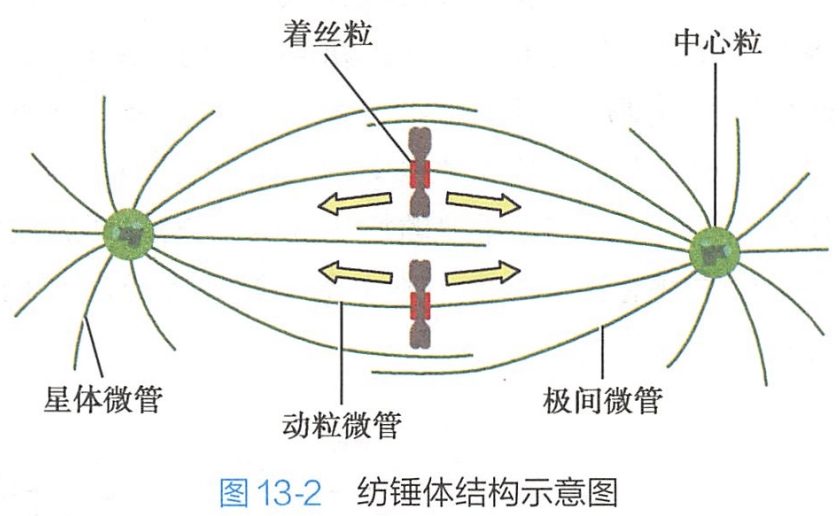
中心体是与染色体分离相关的细胞器,每一中心体由一对中心粒(centriole)及周围无定型基质所构成,这些无定型基质中包含微管蛋白、微管结合蛋白、马达蛋白以及一些与细胞周期调控有关的蛋白质。
中心体是细胞的微管组织中心之一,其周围放射状分布着大量微管,这些微管与中心体一起被合称为星体(aster),星体周围微管在细胞分裂中发挥重要的动力学作用,可分为三类:
- 极微管:极微管为两个星体之间在赤道附近重叠的微管,重叠区微管在动力蛋白的作用下相互滑动,促成星体向两级移动;
- 动粒微管:动粒微管从中心体发生,另一端与染色体动粒结合,其主要作用是通过动粒微管缩短而将染色单体拉向两极;
- 星体微管:星体微管位于星体周围,游离端伸向胞质。经细胞分裂间期已复制后的中心体完全分裂为两个,形成两个星体,星体中的马达蛋白以星体微管作为轨道,利用ATP水解提供的能量沿微管移动,牵引两个子中心体彼此分离,移向细胞的两极,最终两个星体以各自的中心体为两极形成纺锤体。
纺锤体(spindle)是在分裂期出现的特化的亚细胞结构,是一种临时性的梭形细胞骨架结构,由星体微管、极微管和动粒微管纵向排列组成,由中心体作为两极,因状如纺锤而得名(图13-2)。(其实两个星体、星体周围微管相加就是纺锤体吧)
(二)分裂前中期细胞核膜崩解
前中期(prometaphase)细胞变化的主要特征为:核膜崩解、完成纺锤体的装配、染色体列队。
1.核纤层降解、促发核膜崩解
在前期末,因核纤层蛋白多肽链的多个位点发生磷酸化,致使核纤层降解,随后核膜破裂,形成许多断片及小泡,分散于胞质中,在核膜重建时,上述小泡将成为新核膜的组分。
2.纺锤体“捕捉”染色体、完成纺锤体装配,形成有丝分裂器
在前期,两个星体向两极移动,形成分裂极。在驱动蛋白以及其他微管结合蛋白的协助下,由星体发出的极微管、其游离端在赤道面处相互交叠或相互搭桥,形成纺锤体的基本架构;而星体微管与细胞质中的细胞骨架相结合,发挥稳定星体位置的作用;动粒微管与染色体主缢痕部位的着丝粒-动粒复合体结合,捕捉染色体。
纺锤体(包括星体和三种星体周围微管)及与之结合的染色体共同构成有丝分裂器(mitotic apparatus)。
3.染色体列队
前中期,纺锤体两极距离较短,赤道面直径较大,与同一条染色体相连的两极动粒微管并不等长,随着动粒微管的不断聚合与解聚,这种牵引作用造成染色体在震荡中向细胞中央赤道面移动。
(三)分裂中期染色体排列在细胞中央的赤道面
中期(metaphase)的主要特点是由于同一条染色体相连的两极动粒微管等长而达到力量平衡,导致所有染色体排列在细胞中央的赤道面上。此期染色体在形态上比其他任何时期都短粗,同时两条姐妹染色单体的臂较易分离,故特别适合于进行染色体数目、结构等细胞遗传学的研究。
(四)分裂后期细胞姐妹染色单体分离
后期(anaphase)细胞变化的主要特征是染色体两姐妹染色单体分离并移向细胞的两极。姐妹染色单体分离的原因主要与染色体着丝粒分裂有关。
姐妹染色单体原先在着丝粒处依靠粘连蛋白相连,后期粘连蛋白复合体被蛋白酶剪切而崩解,粘合力减小、消失,导致两侧动粒微管对染色单体的拉力与黏连蛋白粘合力的平衡打破,两边的拉力占上风,于是姐妹染色单体分开,而分离后的姐妹染色单体各自成为一个独立的染色体,即子代染色体。
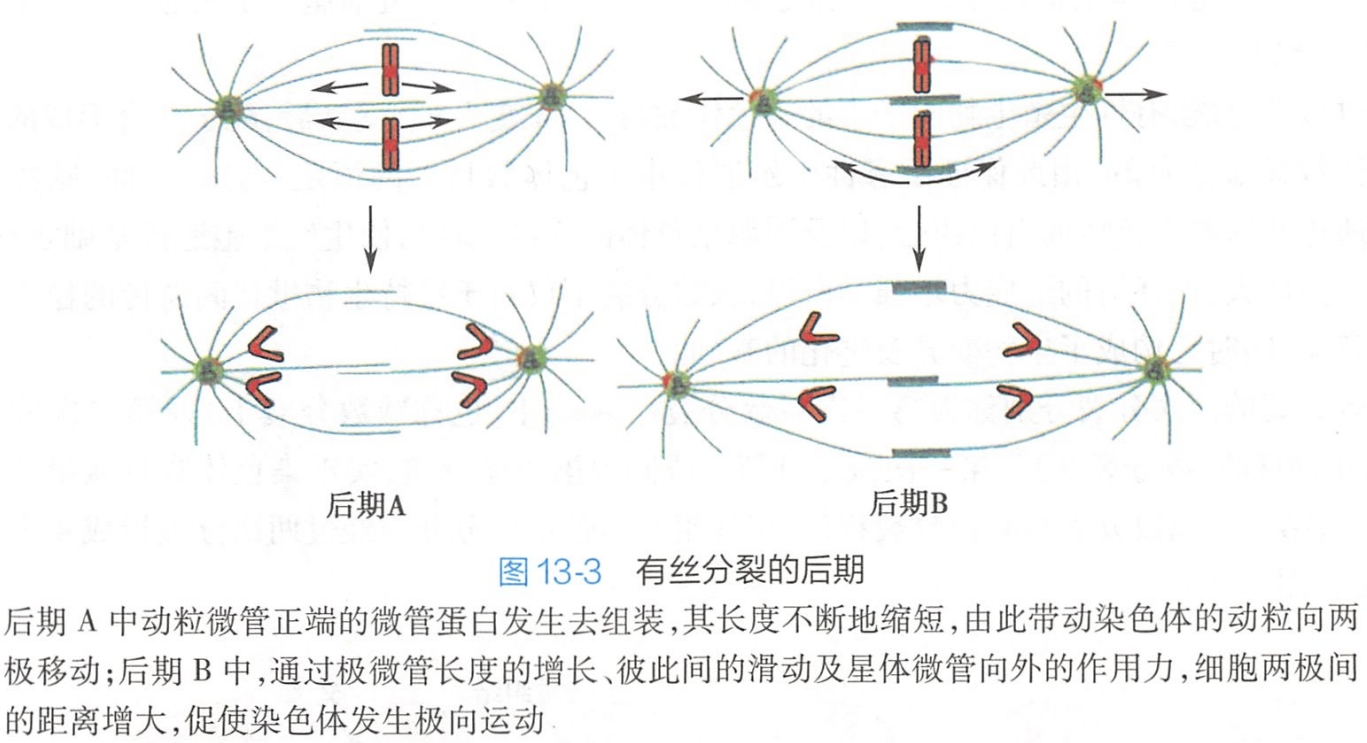
分离后形成的子代染色体以基本相同的速度向两极移动。这种移动是由纺锤体的不同微管参与的两种机制共同作用下实现的。由此可将后期分为后期A和后期B。
- 后期A主要由动粒微管变化介导的。动粒微管解聚而缩短,促发子代染色体向两极移动。
- 后期B主要由极微管变化介导的。在后期A的基础上,极微管聚合而伸长,通过重叠部分微管的长度的增长及彼此间的滑动,同时伴随星体微管向外的作用力,共同促成纺锤体逐步拉长,进一步促进子代染色体移向两极(图13-3)。
(五)分裂末期细胞实现核分裂
末期(telophase)细胞主要的特点是子代细胞的核重建。随着后期末染色体移动到两极,染色体被平均分配的完成,发生了和分裂前期相反的染色体解聚的过程。两套子代染色体分别到达纺锤体两极,动粒微管消失,核膜重新形成,染色体去浓缩恢复间期染色质形态,核仁重新出现。至此,两个子代细胞的核形成,核分裂完成。
(六)依靠收缩环实现胞质分裂
胞质分裂(cytokinesis)是指母细胞胞体一分为二的过程。一些多核细胞(如破骨细胞、骨骼肌细胞和肝细胞)只发生核分裂而无胞质分裂,因而成为多核细胞。但是在典型的有丝分裂中,胞质分裂伴随每次核分裂发生。胞质分裂通常开始于有丝分裂后期,完成于末期。
1.收缩环实现胞质分裂
当细胞分裂进入后期末或末期初,在中部质膜的下方,出现了由大量肌动蛋白和肌球蛋白聚集形成的环状结构,即收缩环(contractile ring)。
此时胞质中的纺锤体也逐渐解体,残存的微管及一些囊泡也聚集于子代细胞核之间的细胞中部,所形成的环形致密层称为中体。 #?
由收缩环中肌动蛋白、肌球蛋白装配而成的微丝束,通过相互滑动使收缩环不断缢缩,直径减小,与其相连的细胞膜逐渐内陷,形成分裂沟,细胞表面出现皱褶;随着分裂沟不断加深,细胞逐步凹陷,当分裂沟深至中央时,一方面细胞在此发生断裂,由收缩环完成胞质分裂,另一方面通过细胞内小泡融合插入收缩环邻近的细胞膜,用于补充细胞膜,保证新分裂形成的细胞与亲代细胞具有相似的表面积。
现已知道,分裂沟发生的时间及部位与纺锤体的位置密切相关,纺锤体的位置决定着两个子代细胞的大小,当纺锤体处于细胞中央时,细胞对称分裂,产生的两个子代细胞大小均等,成分相同。相反,不在细胞中央的纺锤体将导致细胞不均等分裂,所产生的子代细胞在大小与成分上均有差异。
2.细胞器的非绝对均等分配
每个子细胞必须得到母细胞中基本的细胞成分,包括各种膜性细胞器。当细胞进入有丝分裂后期,细胞器的数目或体积都大致扩增一倍,如线粒体通常其数量可在每个细胞周期中简单加倍、内质网和高尔基复合体则体积加大等,在胞质分裂阶段,各种细胞器较为均匀,但非数量绝对均等地分配到子细胞。
二、减数分裂

减数分裂(meiosis)是生殖细胞形成过程中的特殊有丝分裂。减数分裂的主要特征是DNA只复制一次,而细胞连续分裂两次,产生四个子代细胞,每个子代细胞中染色体数目比亲代细胞减少一半,成为仅具单倍体遗传物质的配子细胞。由于减数分裂只发生于生殖细胞的成熟阶段,因此又称为(性)成熟分裂。
经过减数分裂,有性生殖生物配子中的染色体数目由2n变为n。受精后,配子融合形成的受精卵中染色体数又恢复为2n,由此保证了有性生殖遗传中染色体数目上的恒定。
另一方面,减数分裂过程中可通过非同源染色体的自由组合,以及同源染色体的交换、重组,使生殖细胞遗传基础多样化,生物后代变异增大,对环境的适应力增强。与有丝分裂相比,同源染色体的配对是减数分裂的显著特征。配对导致母方染色体上某一片段与同源的父方染色单体相应的片段发生互换。在染色体交叉互换过程中,母方的染色单体和同源的父方的染色单体上DNA的双螺旋结构都是打开的,这有利于在两条非姊妹染色单体间进行某一片段的相互交换,这一过程就是遗传重组。这种同源非姊妹染色单体间的交叉互换增加了后代细胞中的基因类型。
在减数分裂过程中,性染色体也要配对。雌性哺乳动物有两条X染色体,它们能够像其他同源染色体那样配对。但是雄性个体有一条X染色体和一条Y染色体,它们不是同源染色体。有证据表明,在减数分裂前期I也能发生X染色体与Y染色体同源区域的配对及交叉互换,因为在X染色体和Y染色体的末端存在一个小的区域,两者的这个区域是具有同源性的。这种配对、交叉保证X和Y染色体可以连接在纺锤体上,便于染色体分离,结果只产生两种类型的精子,即含有X染色体的精子和含有Y染色体的精子。
所以,减数分裂不仅对于维持生物世代间遗传的稳定性具有重要的意义,同时也构成了生物变异及进化的基础。
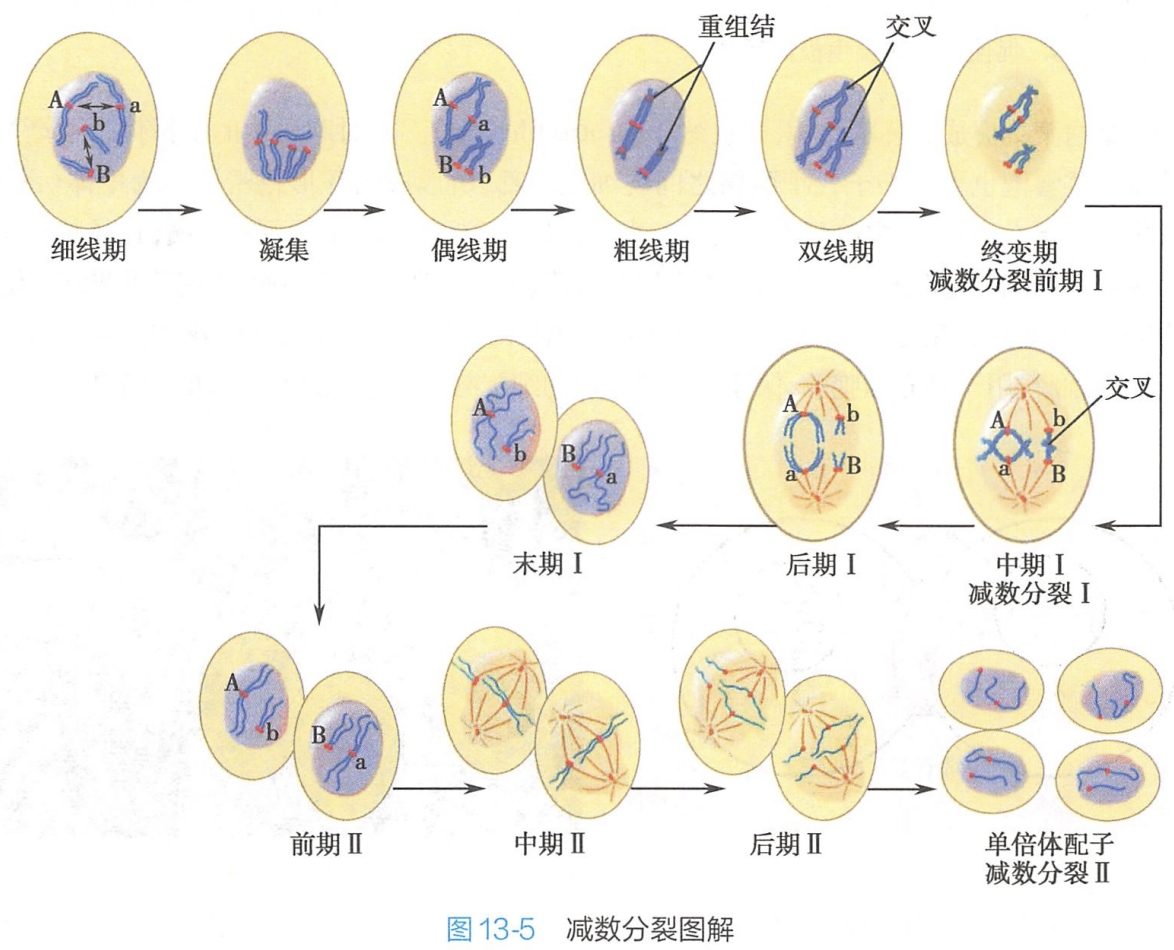
减数分裂的两次分裂分别称为第一次减数分裂(meiosis Ⅰ,也称减数分裂Ⅰ)及第二次减数分裂(meiosisⅡ,也称减数分裂Ⅱ)。第一次减数分裂完成同源染色体分离,实现染色体数目减半及遗传物质的交换;第二次减数分裂与有丝分裂相似,实现姐妹染色单体分开。经过两次分裂形成4个单倍体子代细胞(图13-5)。
(一)第一次减数分裂进程中细胞内发生复杂的生化和形态变化
第一次减数分裂可进一步分为前期Ⅰ、中期Ⅰ、后期Ⅰ和末期Ⅰ。
1.前期Ⅰ
减数分裂的特殊过程主要发生于前期Ⅰ。其主要事件为同源染色体配对、交换与重组,随后随机分离进入两个子代细胞。通常将前期Ⅰ人为划分为5个时期:细线期、偶线期、粗线期、双线期、终变期。
(1)细线期(leptotene stage):
此期细胞中,核及核仁的体积均增大,在分裂间期已经完成复制的染色质开始凝集,每一染色体均具有两条染色单体,但在光镜下仍呈单条细线状,染色单体的臂未完全分离,这可能因为染色体上某些DNA片段的复制尚未完成。
细线状染色体通过其端粒附着于核膜上,在局部出现成串的、大小不一的珠状结构,称为染色粒。
(2)偶线期(zygotene stage):
染色质进一步凝集,分别来自父母的、形态及大小相同的同源染色体(homologous chromosome)相互靠近、配对,称为联会(synapsis)。
染色体配对从端粒处开始,同源染色体间出现若干不同部位的接触点,随后这种结合沿其长轴迅速扩展,直至同源染色体侧面紧密联会。同源染色体完全配对后形成的复合结构即为二价体(bivalent),因其共有四条染色单体,又被称为四分体(tetrad)。
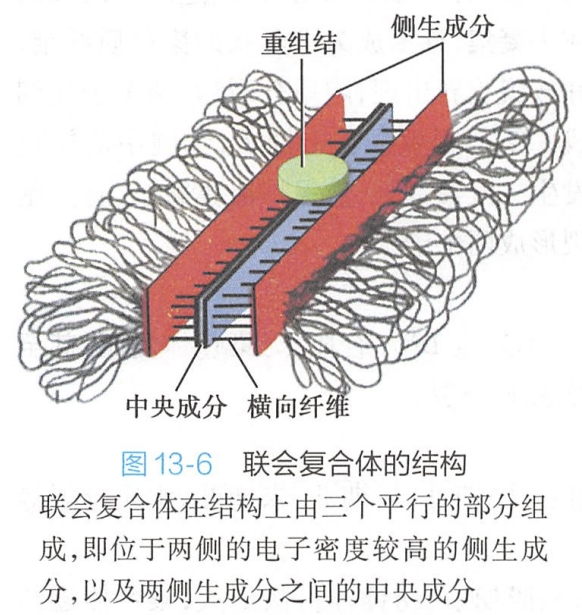
在联会的同源染色体之间,沿纵轴方向形成了一种特殊的结构,称联会复合体(synaptonemal complex,SC),在电镜下它包括三个平行的部分:侧生成分宽约20~40nm,位于复合体两侧,电子密度较高,其外侧为同源染色体DNA。两侧生成分之间电子密度较低的区域为中间区,宽约100nm,其中央为电子密集的中央成分,宽约30m。侧生成分与中央成分之间存在横向排列的纤维,三者大致呈直角相连,每个横向纤维之间距离约20~30nm,因而使联会复合体像一条“拉链”将同源染色体连接在一起(图13-6)。
联会复合体主要由蛋白质、RNA及少量DNA组成。联会复合体是同源染色体配对过程中细胞临时生成的特殊结构,对于同源染色体配对、交换与分离均发挥重要作用。联会复合体的形成中还可能有DNA的参与,偶线期细胞中存在0.3%的DNA合成,称为Z-DNA (好像不一样),如抑制其合成,联会复合体的组装将受到阻止。
联会复合体的形成对于稳定二价体中同源染色体紧密的配对有重要意义。在联会起始阶段,当染色质凝集程度较低时,同源染色体之间主要通过其特定位点上碱基间的互补进行接触。
当联会复合体随着染色质进一步的凝集逐渐形成,其组成中加入了某些单链DNA序列以及蛋白质序列识别分子,由此构成一个有组织的网络,从而促使了同源染色体之间的配对进一步发展,直至完成。
(3)粗线期(pachytene stage):
粗线期持续时间较长,可达几天,甚至几月。通过联会紧密结合在一起的两条同源染色体,进一步地凝集而缩短、变粗,同源染色体间出现染色体段的交换及重组,因此,该期又称为重组期。
联会复合体中央出现一些椭圆形或球形、富含蛋白质及酶的棒状结构,称为重组结(recombination nodule)。多个重组结相间地分布于联会复合体上,将同源非姐妹染色单体的DNA相对区域结合在一起,发生活跃的DNA片段交换事件,导致基因重组。
在粗线期,除合成减数分裂期特有的组蛋白外,同时合成少部分DNA,称为P-DNA,主要编码与DNA剪切和修复相关的酶,在重组过程中发挥DNA修复等作用。
(4)双线期(diplotene stage)
双线期持续时间长短变化较大,一般持续时间较长。作为临时性亚细胞结构的联会复合体在双线期发生去组装,逐渐趋于消失,紧密配对的同源染色体相互分离。同源染色体的大部分片段分开,但仍在非姐妹染色单体之间的某些部位上,残留一些接触点,称为交叉(chiasma)(图13-7)。
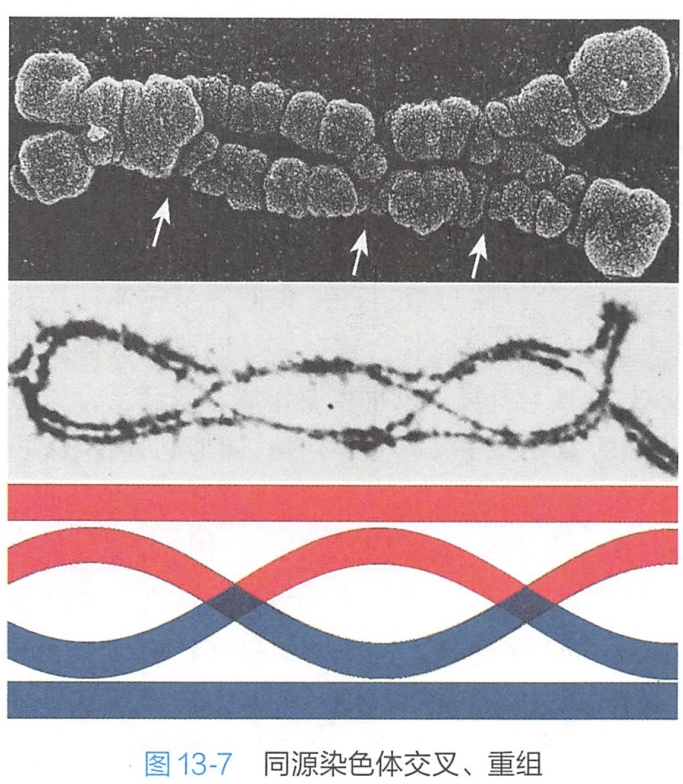
交叉被认为是粗线期同源染色体交换的形态学证据。一般每个染色体至少有一个交叉;染色体较长,交叉也较多。人类平均每对染色体的交叉数为2~3个。随着双线期的进行,交叉点逐渐移向染色体两端,数目也由此减少,这种现象称为交叉端化(chiasma terminalization)。
(5)终变期(diakinesis stage)
同源染色体进一步凝集,核仁消失,交叉端化继续进行。终变期末,同源染色体仅在其端部靠交叉结合在一起,同源染色体重组完成。核膜逐渐解体,纺锤体装配完成,在其作用下染色体开始移向细胞中部的赤道面上。终变期结束标志着前期Ⅰ完成。
2.中期I
以端化的交叉连接在一起的同源染色体即四分体,向细胞中部汇集,最终排列于细胞的赤道面上,通过动粒微管分别与细胞不同极相连,每个二价体的两个动粒分别位于赤道面的两侧,各自面向相对两极,而一侧纺锤体动粒微管只连接于同侧的动粒上,由此决定二价体中每条染色体的后续的相反去向。 #? 一个染色体有两个单体,应该有俩动粒结构域,为啥第一次减数分裂就是共有一个,第二次减数分裂就有俩
3.后期I
由于每个同源染色体的两条姐妹染色单体共有一个着丝粒和动粒,受纺锤体微管的牵拉作用,同源染色体彼此分离,而姐妹染色单体并不分开,包含两条姐妹染色单体的同源染色体开始分别移向细胞的两极,结果导致每极的染色体数为细胞原有染色体数的一半,所以,后期I是减数分裂中染色体减半的关键时期。
同时,同源染色体向两极的移动是随机的,因此,非同源染色体之间以自由组合的方式进入两极,有利于生物变异与进化。
4.末期I
细胞在末期I存在两种类型的变化:
- 类似有丝分裂末期,在到达细胞两极的染色体去凝集,逐渐成为细丝状的染色质纤维,核仁和核膜重新出现,胞质分裂后,两个子代细胞形成。
- 某些生物在末期I,细胞中的染色体不发生去凝集,而依然保持凝集状态,直至胞质分裂形成两个子细胞。
(二)第一次减数分裂后出现短暂的间期
与有丝分裂间期相比,减数分裂间期通常持续时间较短,不发生DNA合成,无染色体复制,甚至某些生物没有间期,第一次减数分裂结束后,直接进入第二次减数分裂。
(三)第二次减数分裂与有丝分裂过程相似
第二次减数分裂过程与有丝分裂基本相同,可分为前期Ⅱ、中期Ⅱ、后期Ⅱ、末期Ⅱ、胞质分裂等5个时期。
1.前期Ⅱ
末期I松散的染色体重新凝聚,核仁消失,核膜崩解,纺锤体再次形成,染色体逐渐向细胞中央的赤道面移动。
2.中期Ⅱ
染色体整齐排列在赤道面,两个姐妹染色单体分别通过各自的动粒与动粒微管相连,朝向纺锤体两极。
3.后期Ⅱ
姐妹染色单体分离,移向两极。
4.末期Ⅱ与胞质分裂
染色体去凝集,成为染色质纤维,核仁和核膜重新出现,经胞质分裂,新的子代细胞形成,子细胞是染色体数目为n的单倍体细胞。
三、无丝分裂
无丝分裂(amitosis)又称为直接分裂(direct division)。
无丝分裂过程中,间期细胞核经复制后直接分裂成大小基本相等的两部分,期间不形成染色体和纺锤体,核膜也不消失,由亲代细胞直接断裂形成子代细胞,因此,两个子代细胞所获得的遗传物质和其他胞质成分并不一定是均等的,无丝分裂中细胞维持其遗传稳定性的机制目前仍不清楚。
无丝分裂是低等生物细胞增殖的主要方式,但也存在于高等生物的组织细胞,例如动物的上皮组织、疏松结缔组织、肌组织及肝脏等细胞。另外,创伤、癌变及衰老的细胞中也能进行无丝分裂。
无丝分裂具有能量消耗少、分裂迅速、分裂中细胞仍可执行其功能等特点,其快速性与便捷性有利于细胞应激并适应外界环境变化。