细胞核
真核细胞中,除哺乳动物的成熟红细胞和高等植物韧皮部的成熟筛管等少数细胞之外,都含有细胞核。
常用细胞核与细胞质的体积比,即核质比(nuclear-cytoplasmic ratio)来表示细胞核的相对大小。
大多数细胞为单核,但也有双核和多核的,如肝细胞、肾小管细胞和软骨细胞有双核,而破骨细胞的核可达几百个。细胞核通常位于细胞的中央,但也可因细胞中分泌颗粒的形成或包含物的推挤而发生位移。在含有分泌颗粒的腺细胞中,核多偏于细胞的一端。
分裂间期的细胞核称为间期核,只有在间期才能看到完整的细胞核。间期核由核膜、染色质、核仁和核基质(核骨架)等构成(图8-1)。
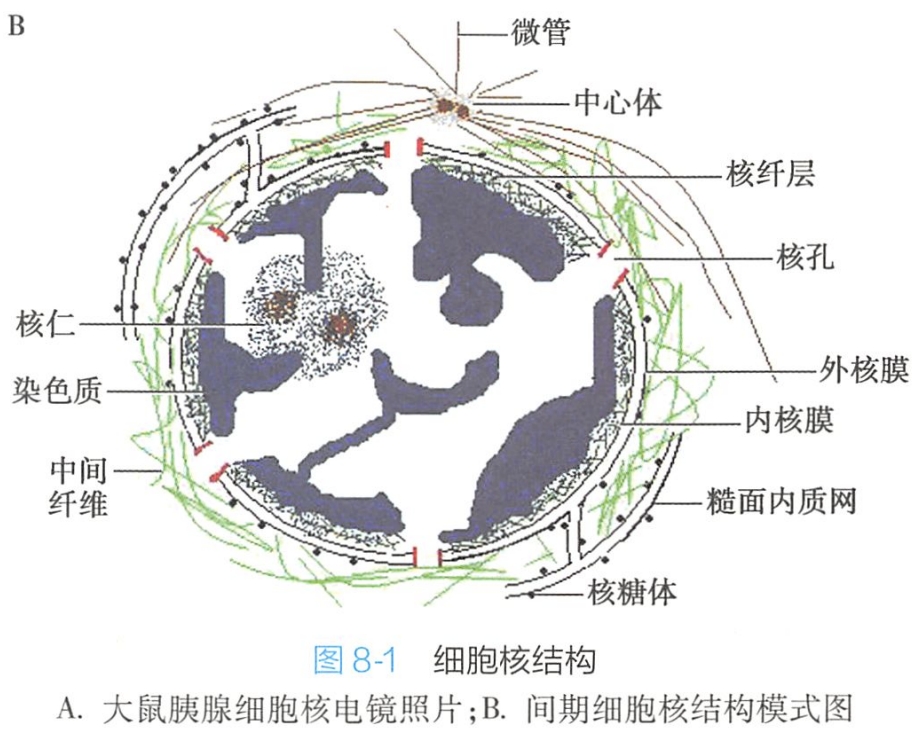
核膜
核膜(nuclear membrane)又称核被膜(nuclear envelope),是细胞核与细胞质之间的界膜。它将细胞分成核与质两大结构与功能区域:DNA复制、RNA转录与加工在核内进行,蛋白质翻译则在细胞质中进行。
一、核膜的化学组成
核膜的主要化学成分是蛋白质和脂类,此外,还有少量核酸成分。
1.蛋白质
核膜中的蛋白质约占65%~75%,包括组蛋白、基因调节蛋白、DNA和RNA聚合酶、RNA酶等。
2.酶
核膜所含的酶类与内质网极为相似,如内质网的标记酶G6PD也存在于核膜。与电子传递有关的酶类,均见于内质网及核膜上,但其含量有差异,如内质网上细胞色素P450的含量高于核膜。
3.脂类
核膜中所含的脂类也与内质网相似,都含有卵磷脂和磷脂酰乙醇胺,以及胆固醇、甘油三酯等。但其含量有差别,核膜所含的不饱和脂肪酸的含量都较低,而胆固醇和甘油三酯的含量却较高。
这种结构成分的相似性和特异性,不仅说明核膜与内质网的密切关系,同时也说明两者也具有各自的结构特点。
二、核膜的结构
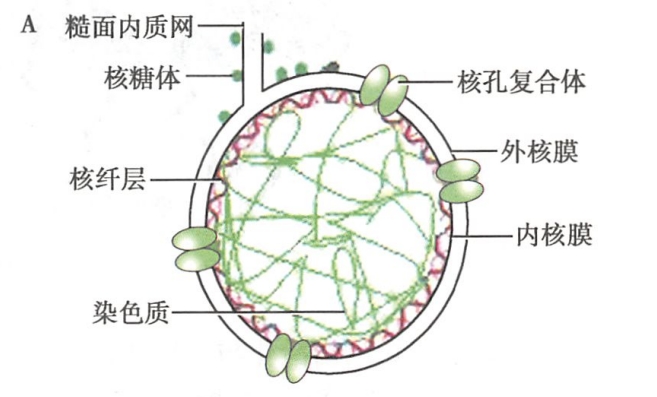
在电镜下,核膜是由内外层核膜、核周隙、核孔复合体和核纤层等结构组成(图8-2)。
因内、外核膜的组成成分和结构都有差异,因此核膜是一种不对称的双层膜结构。
(一)外核膜与糙面内质网相连续
外核膜(outer nuclear membrane)为核膜中面向胞质的一层膜,在形态和生化性质上与细胞质中的糙面内质网膜相近,并与糙面内质网相连续。外核膜外表面有核糖体附着,可进行蛋白质的合成。
外核膜与细胞质相邻的表面可见中间纤维、微管形成的细胞骨架网络,这些结构的存在起着固定细胞核并维持细胞核形态的作用。
(二)内核膜表面光滑包围核质
内核膜(inner nuclear membrane)与外核膜平行排列,表面光滑,无核糖体附着,核质面附着一层结构致密的纤维蛋白网络,称为核纤层,对核膜起支持作用。
(三)核周隙为内、外核膜之间的缓冲区
内、外层核膜在核孔的位置互相融合,两层核膜之间的腔隙称为核周隙(perinuclear space)。核周隙与糙面内质网腔相通,内含有多种蛋白质和酶类。
(四)核孔复合体是由多种蛋白质构成的复合结构
在内外核膜的融合之处形成环状开口,称为核孔(nuclear pore)。
核孔的数目、疏密程度和分布形式随细胞种类和生理状态不同而有很大的变化,一般来说,动物细胞的核孔数多于植物细胞;代谢不活跃的细胞中核孔数较少,例如晚期有核红细胞与淋巴细胞的核孔数为1~3个/m;但在RNA转运速度高,蛋白质合成旺盛的细胞中核孔数目较多,例如肝、肾等高度分化但代谢活跃的细胞。
核孔并非单纯由内外两层核膜融合形成的简单孔洞,而是由多种蛋白质以特定方式排列形成的复合结构,称为核孔复合体(nuclear pore complex,NPC)(图8-3)。
目前比较普遍被接受的是捕鱼笼式(fish-trap)核孔复合体模型(图8-2)。该模型认为核孔复合体的基本结构包括以下四个部分:
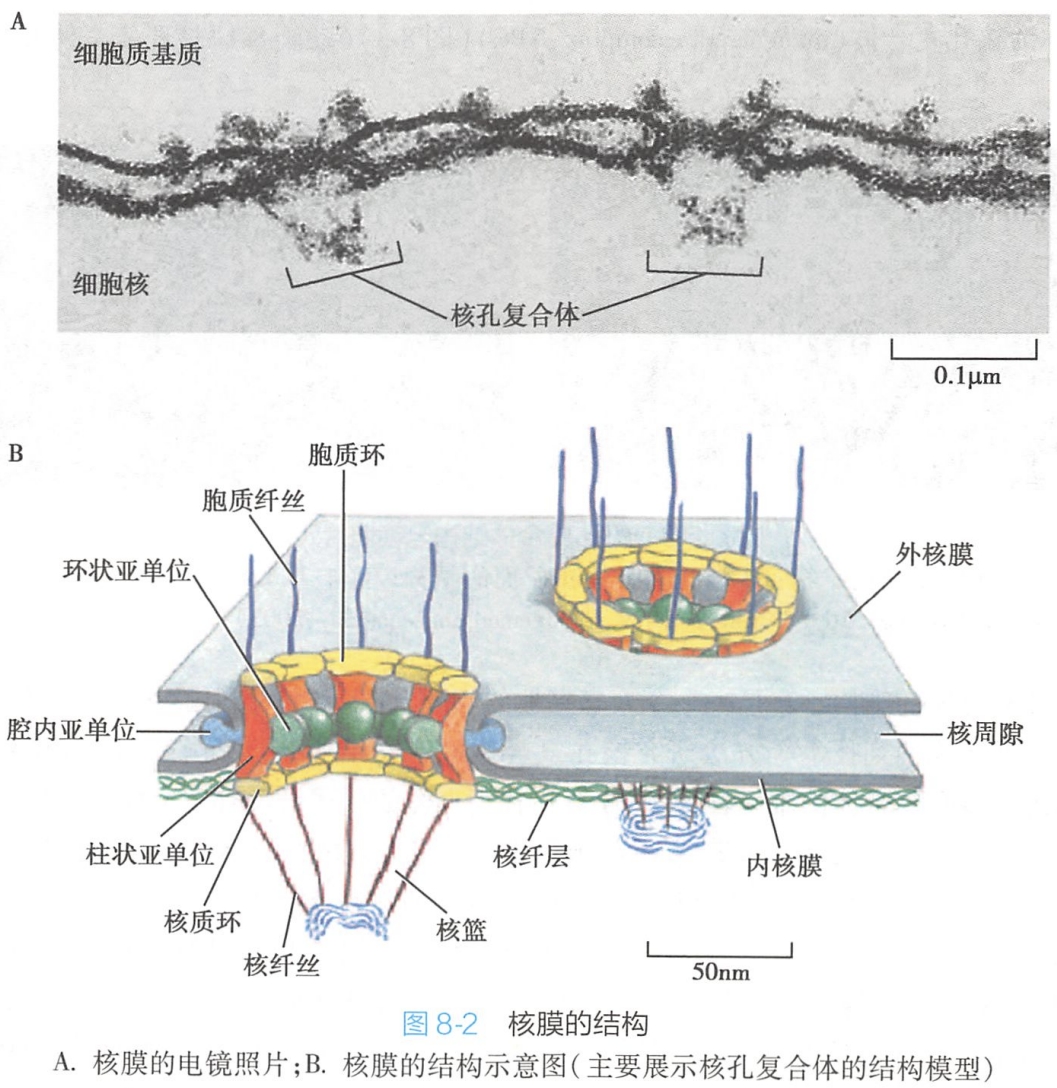
- 胞质环(cytoplasmic ring):位于核孔复合体结构边缘胞质面一侧的环状结构,与柱状亚单位相连,环上对称分布8条短纤维,并伸向细胞质;
- 核质环(nucleoplasmic ring):位于核孔复合体结构边缘核质面一侧的孔环状结构,与柱状亚单位相连,在环上也对称分布8条纤维伸向核内,纤维末端形成一个由8个颗粒组成的直径约60nm的小环,构成捕鱼笼似的结构,称核篮(nuclear basket);
- 辐(spoke):是由核孔边缘伸向中心的结构,呈辐射状八重对称分布,把胞质环、核质环和中央栓连接在一起。辐的结构较复杂,可进一步分为三个结构域:
- 柱状亚单位(column subunit),位于核孔边缘,连接胞质环与核质环,起支撑作用;
- 腔内亚单位(luminal subunit),位于柱状亚单位外侧,与核膜的核孔区域接触,穿过核膜伸入双层核膜的核周隙,起锚定核孔复合体的作用;
- 环状亚单位(annular subunit),位于柱状亚单位之内,是靠近核孔复合体中心部分,由颗粒状结构环绕形成;
- 中央栓(central plug):又称中央颗粒(central granule),位于核孔复合体的中心,是呈颗粒状或棒状的运输蛋白质,其在核质交换中发挥一定的作用。
(五)核纤层是紧贴内核膜的纤维蛋白网
核纤层(nuclear lamina)是位于内核膜内侧与染色质之间的一层由高电子密度纤维蛋白质组成的网络片层结构。在细胞分裂中对核膜的破裂和重建起调节作用。。核纤层的主要化学成分是核纤层蛋白(lamin)。
在哺乳类细胞中,核纤层是由3种属于中间纤维性质的多肽组成,分别称为核纤层蛋白A、B、C。核纤层蛋白具有中间纤维蛋白的α-螺旋区同源的氨基酸顺序。
Lamin A和Lamin C是由同一基因编码的不同的加工产物。两种蛋白质之间仅在一COOH末端不同。核纤层蛋白实际上是一种中间纤维蛋白。核纤层与核膜、核孔复合体及染色质在结构和功能上有密切的联系(图8-5)。
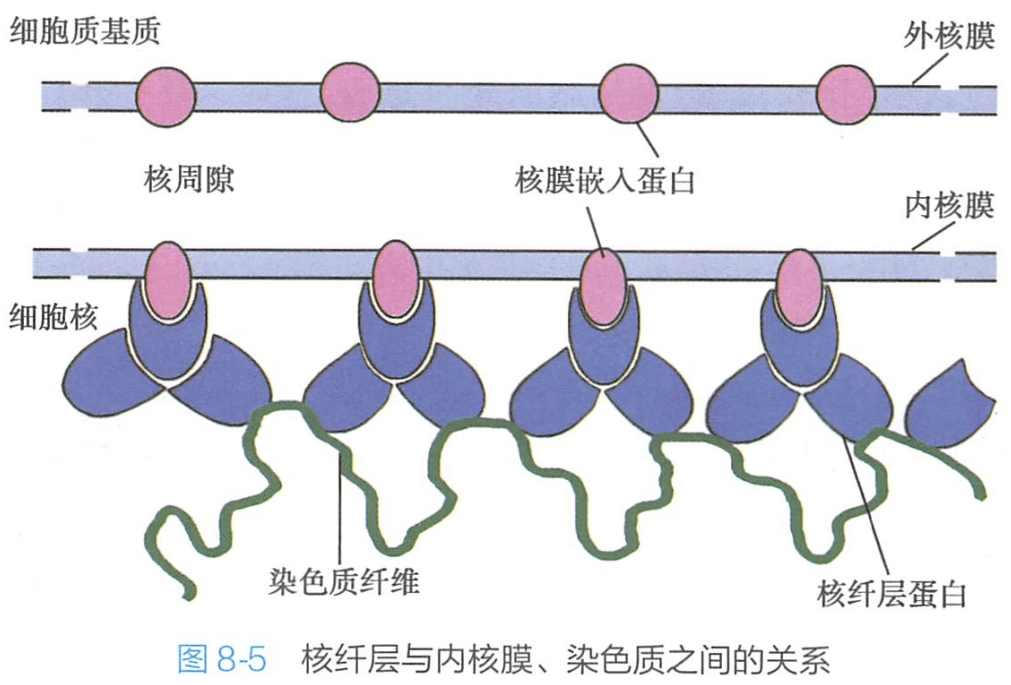
1.核纤层在细胞核中起支架作用
用高盐溶液、非离子去污剂和核酸酶去除大部分核物质,只有核孔复合体与核纤层存留,但仍能维持核的轮廓。此外,核纤层与核骨架及穿过核膜的中间纤维相连,使胞质骨架和核骨架形成一连续网络结构。
2.核纤层与核膜的崩解和重建密切相关
真核细胞在细胞分裂过程中核膜经历崩解与重建的变化,内核膜下的核纤层也经历解聚与聚合的变化。生化分析表明,Lamin A、Lamin B和Lamin C均有亲膜结合作用,其中以Lamin B与核膜的结合力最强。同时,在内核膜上有Lamin B受体,可为 Lamin B提供结合位点,从而把核膜固定在核纤层上。
细胞分裂前期,核纤层蛋白磷酸化,核纤层可逆性去组装,发生解聚,使核膜破裂。此过程中 Lamin A与Lamin C分散到细胞质中,Lamin B因与核膜结合力强,解聚后即与核膜小泡结合,这些小泡在细胞分裂末期是核膜重建的基础。
细胞分裂末期,核纤层蛋白发生去磷酸化,核纤层蛋白又重新在细胞核的周围聚集,核膜再次形成。说明核纤层蛋白的磷酸化与去磷酸化在细胞的有丝分裂过程中与核膜的崩解与重建密切相关。
3.核纤层与染色质凝集成染色体相关
核纤层蛋白可与染色质上的一些特殊位点相结合,为染色质提供了结构支架。
细胞分裂间期,染色质与核纤层紧密结合,因此不能螺旋化成染色体;而在细胞有丝分裂前期,随着核纤层蛋白的解聚,染色质与核纤层蛋白的结合丧失,染色质逐渐凝集成染色体。
4.核纤层参与DNA的复制
重建的没有核纤层的细胞核,虽然细胞核里具有DNA复制过程所需要的蛋白质和酶,但却不能进行DNA的复制,这表明只有染色质而无完整的核膜是不能复制DNA的,提示核纤层参与了DNA的复制。
三、核膜的功能
(一)核膜为基因表达提供了时空隔离屏障
原核细胞因缺乏核膜,遗传物质DNA分子是分布于细胞质中的,RNA转录及蛋白质合成也均发生于细胞质,在RNA3'端转录尚未结束时,其5'端即已被核糖体结合,并开始进行蛋白质的合成,致使RNA转录本在进行翻译以前,因时间及空间的缺乏,不能有效地被剪切、修饰。
真核细胞中,核膜将核物质与细胞质物质限定在各自特定的区域,使DNA复制、RNA转录及蛋白质合成在时空上相互分隔进行,建立了遗传物质稳定的活动环境。真核生物的基因结构复杂,转录后需要经过复杂的加工,所以核膜的出现保证了RNA转录后先进行加工、修饰,才能输入细胞质中,进而指导和参与蛋白质的合成,使遗传信息的表达调控过程更加精确、高效(图8-6)。
(二)核膜参与蛋白质的合成
外核膜的表面附着核糖体,可进行蛋白质的合成。外膜的附着核糖体也能合成少量膜蛋白。
(三)核孔复合体控制着核-质间的物质交换
核孔复合体是被动扩散的亲水通道。
水分子和某些离子,以及一些小分子,如单糖、双糖、氨基酸、核苷和核苷酸等,可以自由扩散,穿梭于核质之间。
但对于绝大多数大分子物质的核-质分配则需要经核孔复合体进行主动运输。主动运输具有高度选择性,其选择性表现在三个方面:
- 核孔复合体的直径大小是可调节的,主动运输的功能直径比被动运输大;
- 核孔复合体的主动运输是信号识别与载体介导的过程,需消耗能量;
- 核孔复合体的主动运输具有双向性,兼有核输入和核输出两种功能。它既能把DNA复制、RNA转录所需的各种酶及组蛋白、核糖体蛋白和核质蛋白等,经核孔复合体运进细胞核;同时又能把细胞核内装配好的核糖体大小亚基和经转录加工后的RNA通过核孔复合体运到细胞质
1.亲核蛋白的核输入
在细胞质中游离核糖体上合成、经核孔转运入细胞核发挥作用的蛋白质称为亲核蛋白(karyo- philic protein),常见的如核糖体蛋白、组蛋白、DNA聚合酶、 RNA聚合酶等。
核质蛋白(nucleoplasmin)是一种亲核蛋白,可同组蛋白 H2A、H2B结合,协助核小体的装配。蛋白质的核输入具有选择性(图8-7)。
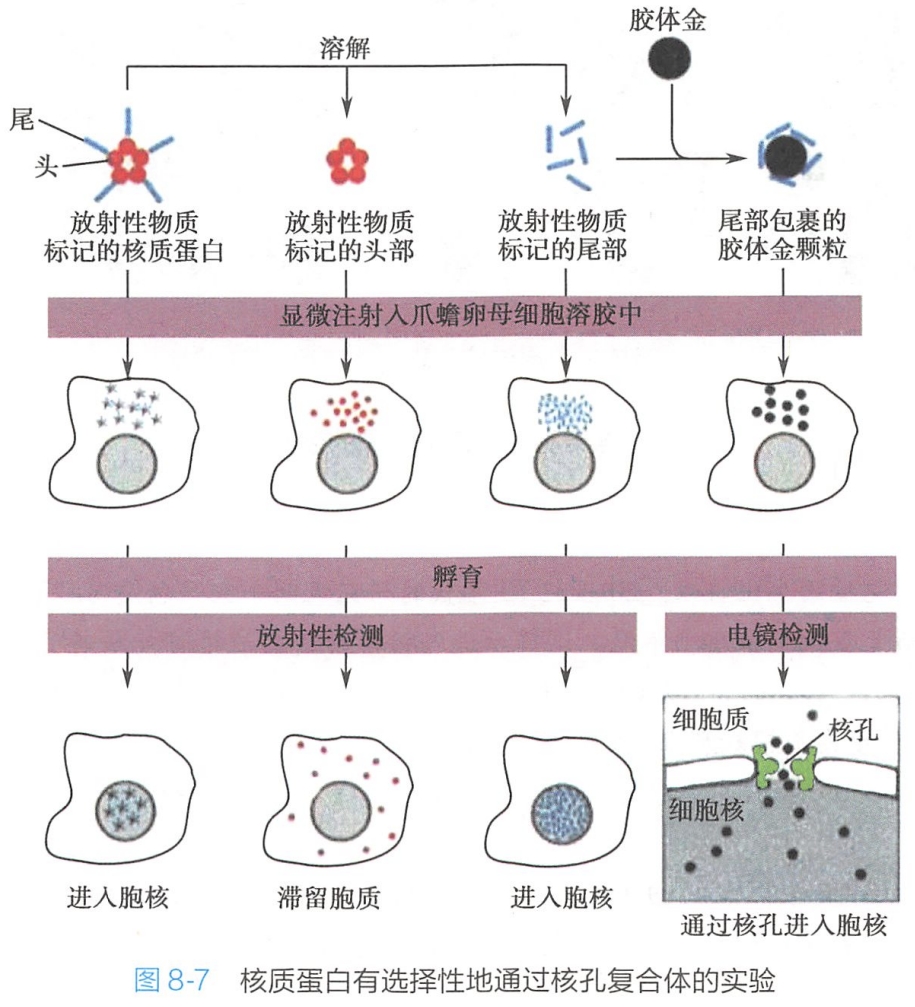
通过对亲核蛋白的序列分析,发现这些亲核蛋白一般都有一段特殊的氨基酸信号序列,正是这些信号序列起到“定向”和“定位”的作用,从而保证蛋白质通过核孔复合体向核内输入,因此,将这一特殊的信号序列命名为核定位序列(nuclear localization sequence,NLS)或核定位信号(nuclear localization signal,NLS)。
具有NLS的蛋白质才能进入核内。通过大量的研究发现NLS是含4~8个氨基酸的短肽序列。不同的亲核蛋白上的NLS不同,但都富含带正电荷的赖氨酸和精氨酸,通常还有脯氨酸。
这些信号序列与指导蛋白质穿膜运输的信号肽不同,NLS可以位于亲核蛋白的任何部位,并且在指导亲核蛋白完成核输入以后不被切除。这个特点有利于细胞分裂完成后,亲核蛋白在子细胞中能够重新输入细胞核。
通过大量研究表明,仅有NLS的蛋白质自身不能通过核孔复合体,它必须通过和NLS受体结合才可通过核孔复合体,这种受体称为输入蛋白(importin)。推测这些输入蛋白在被转运的亲核蛋白与核孔复合体运输装置之间作为一种衔接体蛋白(adaptor protein)而起作用,将在细胞质中结合的蛋白质经核孔复合体转运到核内。
2.RNA及核糖体亚基的核输出
核孔复合体除了把亲核蛋白输入核内以外,还要把新形成的核糖体大、小亚基、RNA和tRNA等输出到细胞质。
核孔复合体除了有亲核蛋白输入信号的受体外,还有识别RNA分子的受体,这些受体称输出蛋白(exportin)。真核细胞中的RNA一般要经过转录后加工,修饰为成熟的RNA分子后才能被转运出核。
核孔复合体对大分子和颗粒物质的运输是双向性的,即将某些物质由细胞质转运入细胞核的同时,也对另一些物质由细胞核向细胞质进行运输。